

 PDF
PDF Citation
Citation Print
Print



Introduction
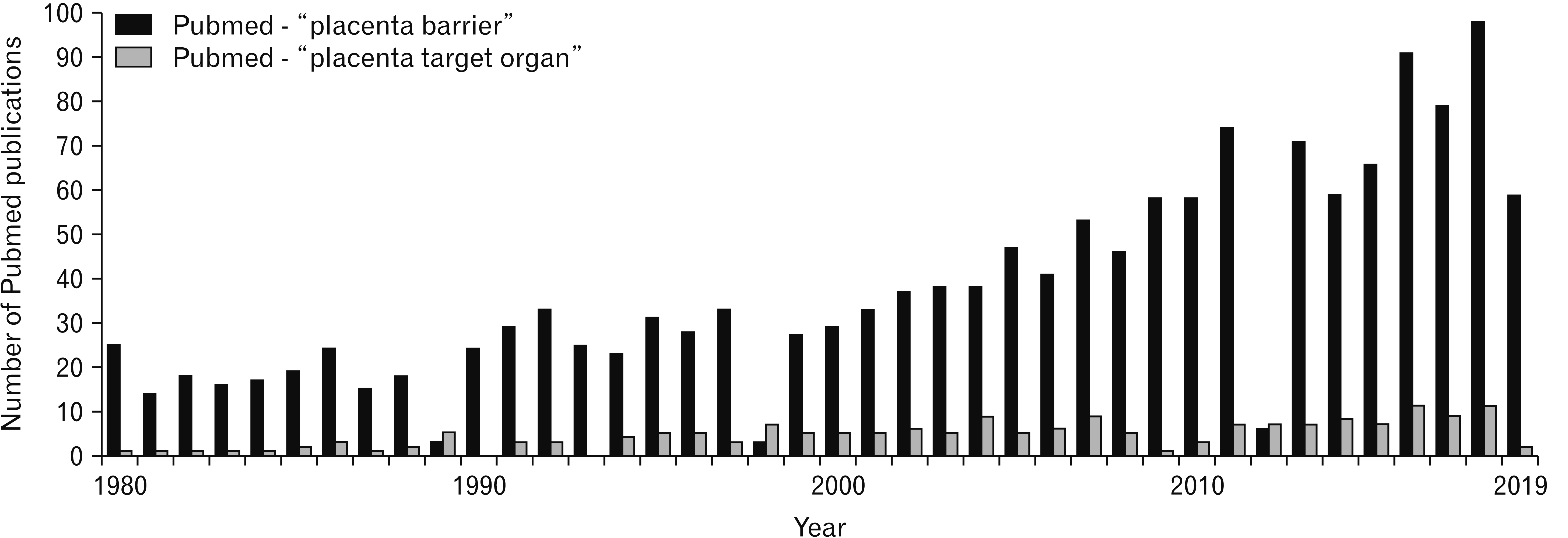
Placenta supports the normal growth and development of the fetus by coordinating gas exchanges, metabolic transfer, immunological functions and by producing, metabolizing and regulating numerous hormones including polypeptide and steroid hormones [1, 2]. Due to the key role of placenta during pregnancy, placental alterations are responsible for many adverse pregnancy outcomes like preeclampsia and intrauterine growth restriction [3-5]. The role of placenta goes beyond pregnancy since placental alterations can be risk factors for cognitive and visual development during childhood [6, 7]. And yet, placenta is more studied as a barrier between mother and fetus in the scientific literature than a target organ for toxic agents (Fig. 1).
Its major role in pregnancy disorders should encourage scientists to consider placenta as a critical organ to further study its response to toxic agents. Hundreds of potential toxic chemicals used in daily life (food, cosmetics, drugs…) and environmental pollutants can be responsible for pregnancy disorders. Placental alterations then represent a growing concern worldwide and placenta toxicology is about to become a fully-fledged toxicology field like cardiotoxicology, hepatotoxicity, neurotoxicity… Placental apoptosis is increased in spontaneous abortion in the first trimester, preeclampsia, post-term pregnancies, and intra-uterine growth restriction [8, 9]. Apoptosis is therefore a key endpoint in the assessment of potential risks for placental toxicity by chemicals.
Placental toxicity is in need of extensive investigation. Several models of placenta are available for toxicology studies. In vivo models are mainly mice, rats and rabbits, but placenta is the organ that shows the greatest diversity across mammalian species in its morphology and tissue organization, mechanisms of implantation and invasion, and endocrine regulation [10-12]. Animal models are consequently not fully suitable for the assessment of toxicological risk in the human placenta. Efforts are besides made to eliminate the use of animals in testing over the past 35 years in accordance to the 3R principle (Reduce, Refine, Replace). Perfused human placenta is the only model that respects the anatomical features of the human placenta [13], but the problem lies in the limited access to whole placenta for obvious technical and logistical reasons. Perfused human placentas as long as placenta-derived primary cultures are therefore not appropriate for high throughput screening. Moreover, it is almost impossible to control the maternal exposition to drugs, chemicals, cosmetics and pollutants that could influence the placental response to the toxic agent being studied. There are several different in vitro approaches currently available to evaluate chemicals toxicity in the human placenta. Immortalized cell lines can be bought in international cell banks like the American Type Culture Collection ATCC, the four main cell lines being HTR-8/SVneo, BeWo, JAR and JEG-3 trophoblastic cells. Contrary to BeWo, JAR and JEG-3 cell lines, the HTR-8/SVneo is not derived from a choriocarcinoma and was immortalized by SV-40 transfection. They show lower expression of proteins expressed by trophoblasts like cytokeratin-7 (CK7) and E-cadherin than BeWo, JEG-3 and JAR cell lines [14]. BeWo and JAR cells are less differentiated than JEG-3 cells. BeWo is best suited for studies on syncytial fusion as they are the only one to be fusigenic. JAR monolayers are unstable, as continued proliferation results in the formation of multilayers where ZO-1 and E-cadherin are lost from the cell surface [15]. With respect to TransEpithelial Electrical Resistance and electrical behaviour, JEG-3 are more similar to primary cells than BeWo and JAR cells [16]. For all these reasons, the human placental JEG-3 cell line is closer to human physiology than the other placenta cell lines and appears to be the best tool for the assessment of chemical toxicity in placenta. According to ATCC’s instructions, JEG-3 cells are grown in MEM culture medium supplemented with 10% fetal bovine serum (FBS) to bring a large number of nutritional and macromolecular factors essential for cell growth and promote cell proliferation. Fetal serum may bind or adsorb chemicals and then mask their cytotoxicity [17, 18]. Low serum medium should be preferred in toxicological studies but how low is the question. We previously published numerous studies using different cell lines (tenocytes, corneal and retinal epithelial cells, macrophages and keratinocytes) that revealed the toxicity of chemicals in culture medium supplemented with serum reduced from 10% to 2.5% [19-22]. Based on our expertise, we suggest that the concentration of serum in JEG-3 cells should be reduced to 2.5% for toxicology studies.
The objective of the present paper is to establish incubation conditions for JEG-3 placental cells to reveal pregnancy disorders induced by chemicals. To achieve this objective, we first studied JEG-3 cells behavior in 2.5% serum compared to 10%, and second, we checked that JEG-3 cells are able to undergo apoptosis in response to chemicals toxic for pregnant women.
Materials and Methods
Materials
All tested chemicals were purchased from Merck (Darmstadt, Germany) except ethanol (VWR Chemicals, Radnor, PA, USA) and perfluorooctanoic acid (PFOA; ThermoFisher Scientific, Waltham, MA, USA).
All cell culture reagents were obtained from Gibco (Paisley, UK). A 96-well microplates were purchased from Corning (Amsterdam, The Nederlands) and Nunc Lab-Tek II Chamber Slide system from Merck.
Antibodies were purchased from Merck (mouse anti-CK7 antibody) and ThermoFisher Scientific (Alexa Fluor 488 goat anti-mouse antibody and isotypic control). Fluorescent probes were obtained from ThermoFisher Scientific.
Fluorescence Resonance Energy Transfer (FRET) and ELISA kits were purchased from Cisbio Biosassays (Codolet, France) and MyBioSource (Vancouver, BC, Canada), respectively.
Cell culture
The choriocarcinoma-derived JEG-3 cell-line (ATCC HTB-36, Manassas, VA, USA), was grown as recommended by ATCC: Minimum Essential Medium Eagle’s medium supplemented with 10% FBS, 2 mM of glutamine, 50 IU/ml of penicillin and 50 IU/ml of streptomycin. Cells were detached using trypsin, counted, and then seeded at 80,000 cells/ml in 96-well microplates (200 μl by well) and Nunc Lab-Tek II Chamber Slide system for immunostaining.
JEG-3 cells behavior in culture medium supplemented with different concentrations of serum
Impact of FBS concentration on cell proliferation
Cells were cultured in three different concentrations of FBS (using the same batch): 0%, 2.5% and 10%. At 24 and 72 hours, cells were detached using trypsin and then counted by the Countess II Automated Cell Counter (ThermoFisher Scientific).
Cell line authentication by Short Tandem Repeat analysis (genetic profile)
Cell line DNA was profiled by Short Tandem Repeat (STR). This technique also checks the lack of cellular cross-contamination [23]. STR analysis was performed by the Human STR Profiling Cell Authentication Service of ATCC.
CK7 immunostaining
The CK7 intermediate filament is an established marker of trophoblastic cells [14, 24]. A 24 hours after seeding in culture medium supplemented with 2.5 or 10% FBS, JEG-3 cells were fixed in 4% paraformaldehyde for 20 minutes, permeabilized in 0.1% Triton X-100 for 10 minutes, saturated with a solution of 1% BSA and 0.1% Tween in PBS for 2 hours, and then incubated overnight at 4°C with mouse anti-CK7 antibody (196 μg/ml) diluted in PBS containing 1% BSA and 0.1% Tween 20. After washing, the cells were incubated with Alexa Fluor 488 goat anti-mouse antibody (4 μg/ml) diluted in PBS containing 1% BSA for 2 hours at room temperature. Nuclei were stained with 300 nM DAPI for 5 minutes and Vectashield (Vector Laboratories, Burlingame, CA, USA) mounting medium was used for microscopy images (EVOS FL, ThermoFisher Scientific). Mouse IgG1 kappa clone P3.6.2.8.1 was used as an isotypic control to help differentiate non-specific background signal from specific antibody signal.
Hormone release quantitation
After 72 hours of incubation in cell culture medium supplemented with 2.5% or 10% FBS, microplates were centrifuged, and cell supernatants were collected. Estradiol was quantified in cell supernatants by FRET technology (HTRF Cisbio Biosassays, Codolet, France) according to manufacturer’s instructions. The detection limit of this assay is 20 pg/ml.
Human placental lactogen (hPl) hormone and human hyperglycosylated Chorionic Gonadotropin (hCG) hormone were measured by sandwich ELISA (MyBioSource) according to manufacturer’s instructions. Sensitivities are <46.875 pg/ml and 39 pg/ml for hPl and hCG dosage, respectively.
Impact of FBS concentration on sodium lauryl sulfate and PFOA cytotoxicity
Cells were incubated with sodium lauryl sulfate or PFOA diluted in culture medium supplemented with either 2.5% or 10% FBS. After 24 hours, cell viability was evaluated using the neutral red assay. Neutral Red solution at 0.4% in water was diluted in culture medium with a ratio of 1:79 to give a final concentration of 50 μg/ml. Neutral Red was distributed in the plates for a 3-hour incubation time at 37°C. The cells were then rinsed with PBS to remove any remaining unincorporated dye. The dye was then released from the cells using a lysis solution (1% acetic acid, 50% ethanol and 49% H2O) and the fluorescence was measured (λex=540 nm, λem=600 nm) using Spark microplate fluorometer (Tecan, Männedorf, Switzerland).
Apoptosis evaluation after incubation with toxic agents
Toxic agents
Tested toxic agents for apoptosis evaluation were detailed in Table 1. Solvents were evaluated alone to discriminate their potential effect (data not shown).
Determination of subcytotoxic concentrations
Known apoptotic agents were diluted in culture medium supplemented with 2.5% FBS and incubated for 24 hours. Before running the apoptosis assay, cell viability was determined using the Alamar blue assay to eliminate necrotic concentrations and only keep subcytotoxic concentrations of the agents. Alamar blue was diluted in culture medium to a working concentration of 9 μg/ml. The cells were incubated with the solution for 6 hours at 37°C. The fluorescence signal was read (λex=535 nm, λem=600 nm) using the Spark cytofluorometer.
Evaluation of chromatin condensation as a hallmark of apoptosis
The UV fluorescent probe Hoechst 33342 enters living and apoptotic cells, intercalating into DNA. The fluorescent signal is proportional to chromatin condensation in apoptosis. The cells were incubated with Hoechst 33342 at 10 μg/ml for 30 minutes at room temperature. The fluorescence signal was read (λex=360 nm, λem=460 nm) using a cytofluorometer (Spark).
Statistical analysis
Means of at least three independent experiments were calculated and normalized to control. A one-way ANOVA followed by Dunnett’s test were performed (α risk=5%) using GraphPad Prism 6 software (San Diego, CA, USA). Thresholds of significance were *P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001 compared to control.
Results
JEG-3 cells behavior in culture medium supplemented with different concentrations of FBS
Impact of FBS concentration on cell proliferation
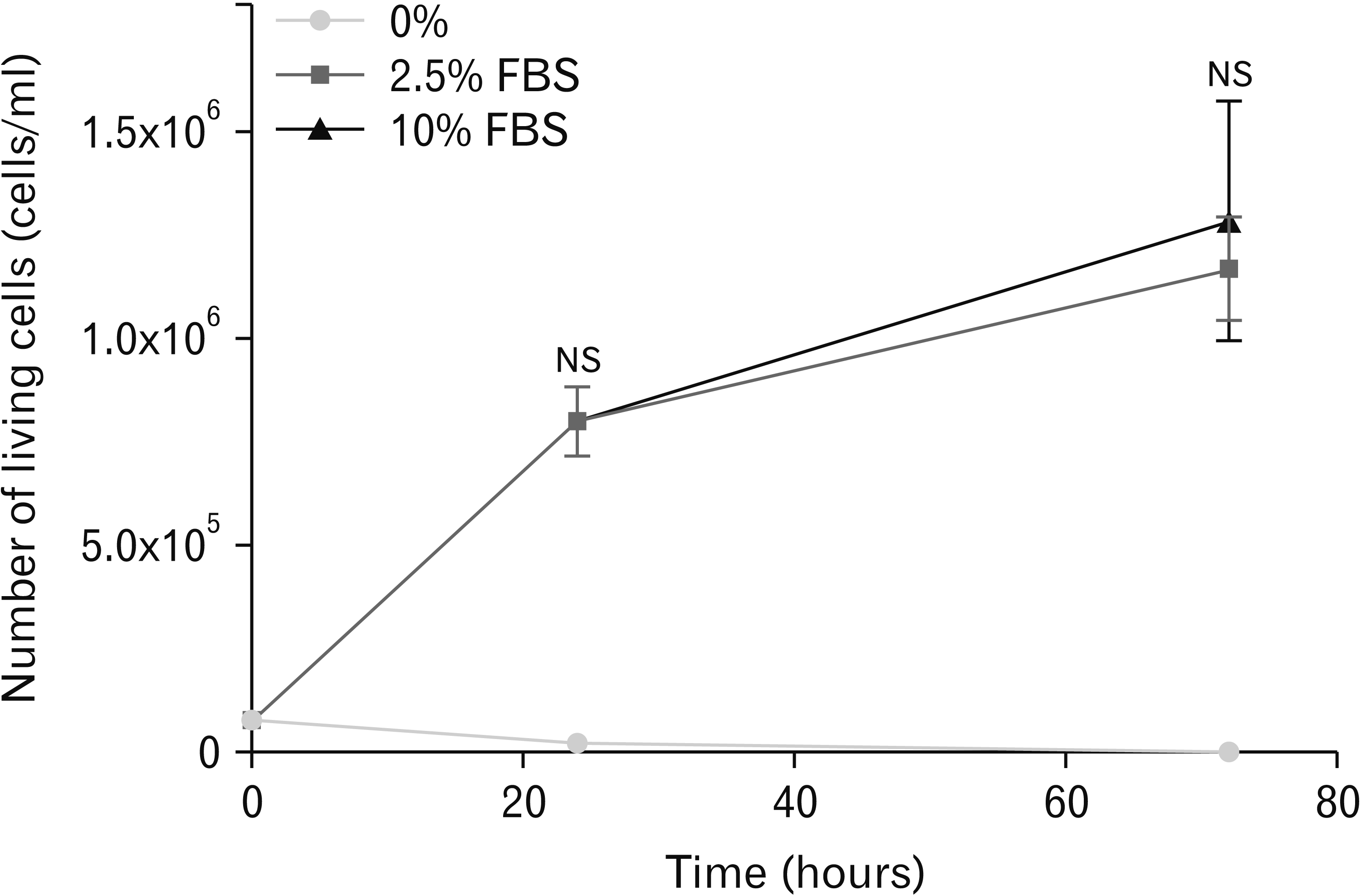
Three percentages of FBS were used: 0%, 2.5%, and 10% in culture medium (Fig. 2).
The percentage of living cells was dramatically decreased after 24 hours in culture medium without FBS (0% FBS); as expected, JEG-3 cells were not able to proliferate without FBS due to a lack of nutritional and macromolecular factors. JEG-3 cell proliferation in culture medium supplemented with 2.5% was similar to proliferation in 10% FBS at 24 and 72 hours.
Cell line authentication by STR analysis
The STR analysis was performed to compare nine STR core markers in JEG-3 cells in culture medium supplemented with 2.5% FBS to JEG-3 cells in 10% FBS (Table 2).
JEG-3 cells in 10% or 2.5% FBS expressed the same STR core markers. Reducing the percentage of FBS in culture medium of JEG-3 cells had no impact on DNA specific loci.
Expression of CK7 as a marker of placental cells
CK7 is a well-known epithelial marker for trophoblast cells and is known to be expressed in JEG-3 cells cultured in 10% FBS [14]. According to our microscopic observations, JEG-3 cells expressed similar levels of CK7 in 2.5% FBS and 10% FBS (Fig. 3A). The expression of CK7 was quantified using ImageJ software (Fig. 3B) and no statistical differences were observed between 2.5% FBS and 10% FBS.
Quantification of estradiol, hyperglycosylated hCG and hPL secretion by JEG-3 cells
We compared the secretion of placental hormones by JEG-3 cells in culture medium supplemented with 10% FBS to JEG-3 cells in 2.5% FBS. After 24 hours in either medium, the levels of each hormone were comparable (Table 3).
Impact of FBS concentration on sodium lauryl sulfate and PFOA cytotoxicity
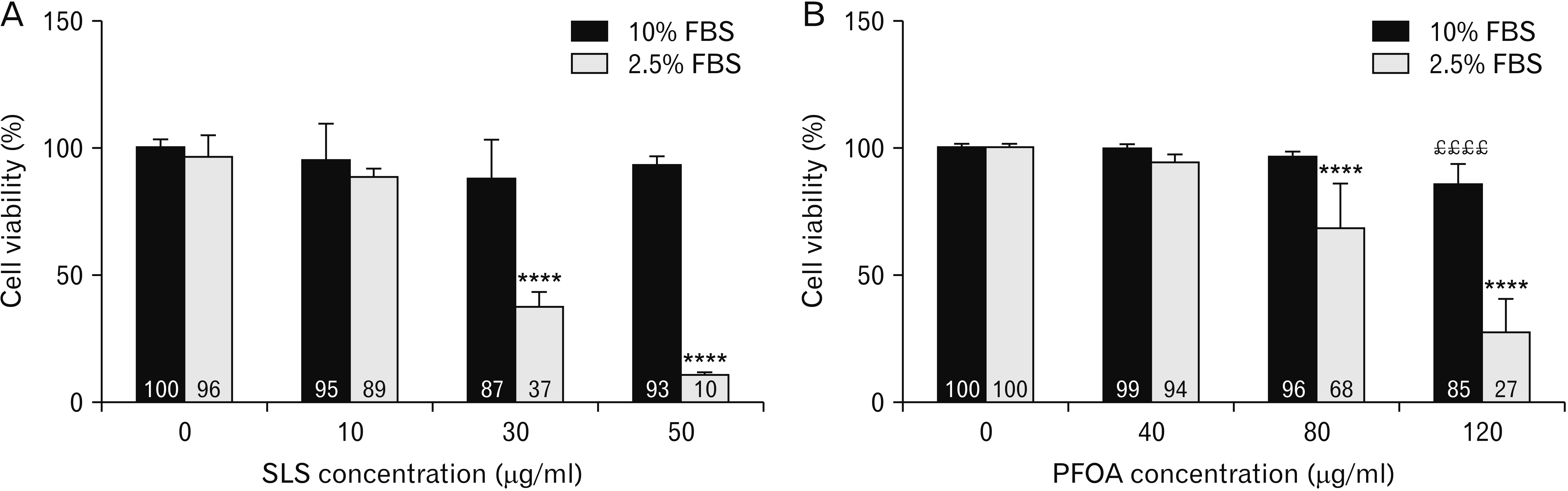
We compared sodium lauryl sulfate (SLS) and PFOA cytotoxicity in 2.5% FBS and 10% FBS, respectively (Fig. 4). At all tested concentrations, SLS diluted in culture medium supplemented with 10% FBS had no effect on JEG-3 cell viability. On the contrary, SLS diluted in culture medium supplemented with 2.5% FBS induced cytotoxicity at 30 μg/ml (37% of living cells, Fig. 4A) and 50 μg/ml (10% of living cells). PFOA cytotoxicity was observed at 80 μg/ml and 120 μg/ml in FBS 2.5% (68% and 27% of living cells, respectively, Fig. 4B) whereas only a slight loss of cell viability was observed at 120 μg/ml in FBS 10% (85% of viable cells). The classic concentration of FBS used for cell culture (10% of total volume) tends to mask SLS and PFOA cytotoxicity contrary to reduced FBS concentration (2.5%).
Based on our results, we pursue our study only using culture medium supplemented with FBS 2.5%; we renamed cells with these incubation conditions JEG-Tox.
Response of JEG-Tox cells to apoptosis inducers
We studied chromatin condensation in JEG-Tox cells after incubation with apoptotic chemicals. Before assessing chromatin condensation, we selected subcytotoxic concentrations i.e. concentrations that result in % of living cells higher than 70 (data not shown). This threshold is recommended in ISO standards and OECD guidelines that assess cytotoxicity on monolayer cells. Subcytotoxic concentrations ranged from 0.1% to 5% for ethanol, from 0.03 to 150 μg/ml for quinalphos, from 2 to 20 μg/ml for bisphenol F, from 0.4 to 16 μg/ml for 4,4’DDT, from 0.1 to 2.5 μg/ml for BAC, from 0.0001% to 0.15% for phenoxyethanol, from 0.2 to 20 μg/ml for propylparaben and from 0.04 to 100 μg/ml for PFOA.
As shown in Fig. 5, all the apoptotic chemicals significantly induced chromatin condensation in JEG-Tox cells. Chromatin condensation was initiated with ethanol 2.5%, quinalphos 0.3 μg/ml, bisphenol F 5 μg/ml, 4,4’DDT 2 μg/ml, BAC 2.5 μg/ml, phenoxyethanol 0.15%, propylparaben 20 μg/ml and PFOA 20 μg/ml; all those concentrations being in accordance with the literature in other cell types [25-31].
Discussion
Chemicals are more concentrated in the placenta than in maternal tissues [32]. Exposure of pregnant women to hazardous chemicals and environmental pollutants like alcohol, pesticides, preservatives, or plasticizers can lead to decreased birth length and weight and increased infant mortality, alterations of developing nervous system and other vital organs, endocrine disruptions [33-36].
Proteins present in FBS can bind chemicals thus masking their potential cytotoxicity and affecting cell response. It was previously proposed that the protein corona formed around particles greatly influences particle toxicity [37]. High FBS concentrations used in growth medium (mainly 10%) are therefore not suitable for toxicity studies. Some of our previous studies on ocular and skin cell lines demonstrated that 2.5% FBS is a good compromise as serum total deprivation induces cell death [38-40]. In this study, we compared placental JEG-3 cells behaviour in 2.5% FBS versus 10% FBS. We first evaluated cell proliferation and observed that JEG-3 cells cultured in 2.5% or 10% FBS have similar proliferation rates, and as expected, cells in 0% FBS did not survive. We second analysed STR core markers and concluded that JEG-3 cells had the same STR core markers and thus the same genotype whether they are cultured in 2.5% or in 10% FBS. We third performed immunochemistry studies to ensure that JEG-3 cells in 2.5% FBS express CK7, a known marker of placental cells. Our results showed that reducing the percentage of FBS in JEG-3 cells does not alter signatures of cell identity such as cell proliferation rate, DNA profile and specific protein expression. JEG-3 cells in 2.5% FBS released similar levels of hCG, hPL, and estradiol to JEG-3 cells in 10% FBS, and thus maintain the endocrine function of human placenta.
In the cytotoxicity study, we didn’t observe any cell death when SLS was diluted in 10% FBS up to 50 μg/ml whereas when it was diluted in 2.5% FBS, SLS induced a dramatic loss of cell viability at 30 μg/ml. Cytotoxicity of PFOA was revealed at 200 μM when it was diluted in 2.5% FBS whereas only a slight loss of cell viability was observed at 300 μM when it was diluted in 10% FBS. It appears that JEG-3 cells in 2.5% FBS are more suitable for toxicological studies than JEG-3 cells in 10% FBS. We renamed JEG-3 cells in 2.5% FBS JEG-Tox cells.
Apoptosis is suggested to be a key mechanism in placental dysfunction. A growing amount of data indeed suggests that uncontrolled placental apoptosis has side effects on both the placenta and maternal physiology [41]. To validate JEG-Tox cells as a pertinent model for the evaluation of placental toxicity, we checked whether they were able to trigger apoptosis after incubation with known apoptotic agents. We selected chemicals that pregnant women can be exposed to such as ethanol through alcohol consumption, preservatives present in cosmetics or drugs, pesticides and cookware coatings. In our experimental conditions, all the tested apoptotic chemicals induced chromatin condensation in JEG-Tox cells.
To conclude, reducing the percentage of FBS from 10%, which is the recommended concentration for cell growth, to 2.5% does not affect neither DNA profile, nor placental marker, nor hormone secretion, but reveals placental toxicity increasing cell sensitivity to chemicals contrary to FBS 10%. JEG-Tox cells can be of great value in placental toxicological studies, especially to study apoptosis that is at the origin of numerous severe pregnancy disorders.
 XML Download
XML Download