

 PDF
PDF Citation
Citation Print
Print



Introduction
Nicotine, the psychoactive component of tobacco products, stimulate midbrain dopaminergic neurons by activating nicotinic acetylcholine receptors on the dopaminergic somata and by balancing synaptic sources of information [1].
Nicotine treatment for 7 days, induced many adverse effects in rats. These included deterioration of memory, neuronal apoptosis, DNA fragmentation, reduced antioxidant enzymes activity and excessive construction of lipid peroxidation and reactive oxygen species [2]. Ceaseless nicotine administration in a high dose (4 mg/kg body weight for 4 weeks in subcutaneous injections), might induce hippocampal and striatal neurodegeneration [3]. Long-term nicotine exposure induced damage of cerebellar white matter in rat model system [4]. Besides, a strong relation has been recognized between tobacco smoke/nicotine and vascular impairment. Several studies have shown that nicotine exposure for short- or long duration results in a negative impact on the function and/or structure of the blood-brain barrier (BBB), which consequently would lead to the risk of brain edema and neuro-inflammation. It is reported that nicotine creates a dose‐dependent change in the BBB permeation of macromolecules [5]; alter cerebrovascular flow [6, 7] alters expression or function of BBB‐associated proteins [8, 9] has deleterious effect on the endothelium; and; increases the permeability of the BBB via a redistribution of tight junction proteins in cerebral micro-vessels [10]. Hawkins et al. [11], 2004 reported that nicotine-induced BBB damage in a mice model.
In a modern study, by using fluorescence microscopy Huang and his coworkers [12] found that nicotine markedly raised expression of S100B (biomarkers for BBB damage) in the hippocampus following use of (1.5 mg/kg/day, of nicotine) (oral delivery) for 3 days (twice per day) [12].
The most prominent function of the pars compacta is motor control; its damage can cause a number of movement disorders including Parkinson’s disease and other dyskinesias. Although many previous studies demonstrated the possible effects of nicotine on striatum functions [13, 14], the impact of nicotine on the substantia nigra compacta (SNc) which is one of the major areas producing dopamine, has been marginally addressed. Accordingly, to address these critical issues, we proposed in this novel study to explore nicotine effect on morphology, and biology of the diverse constituents of the SNc.
Materials and Methods
Chemicals
Nicotine hydrogen tartrate salt (liquid) was purchased from Sigma-Aldrich (St. Louis, MO, USA) and dissolved in normal saline (0.9% sodium chloride) [15].
Animals
Twenty adult male Albino rats (weighing 250-300 g) were obtained from Assiut University Animal House. They were housed under standard conditions (light, temperature, food, and water). Animal care and use was in accordance with procedures outlined in the National Institutes of Health Guidelines. All procedures have been approved by the institutional ethics committee of Assiut University (No. 17200506).
Rats were divided equally into two groups as follows:
Group I (Vehicle-control): 10 animals received normal saline injection (1 ml/kg body weight intra peritoneally) for 11 days.
Group II (Nicotine group): 10 animals were injected subcutaneously with nicotine dissolved in normal saline (0.9 % Na Cl) in a dose of 1.5 mg/kg body weight daily, for 11days [16]. Each rat received a dose of 0.3 mg nicotine in a volume of 0.2 ml solution of saline after adjusting for body weight. Nicotine dose was chosen based on finding that it produces plasma levels of nicotine in rats similar to plasma levels of nicotine found in humans smoking one pack of cigarettes per day. In other word, the dose is close to that from low-to-moderate cigarette smoking [17].
Methods
Biochemical parameters
Nicotine levels: At the end of the experiment, levels of nicotine were detected in the serum according to Massadeh et al. [18] by gas chromatography-mass spectrometry (GC-MS) extraction method (GC-MS apparatus: 7890A-5975B USA-Column DB 5 ms) in Analytical Chemistry Unit-Assiut University. The oven temperature program was initially 100°C for 1 minute, then 10°C/minute to 260°C for 1 minute, with the run time 18 minutes. The oven flow program of 1 ml/minute for 0 minute. The analysis was performed using the selected ion monitoring acquisition mode. The limit of quantitation of nicotine was (1 ng/ml).
Dopamine concentration: It was detected by high-performance liquid chromatography with electrochemical detection (HPLC-ECD). After sacrifice of the animals, the brains were quickly removed, the left and right striata were dissected out, and kept in liquid nitrogen. Striatal tissue was weighed and placed in 0.2 ml ice cold 0.01 mM perchloric acid containing 0.01% EDTA and 60 ng 3, 4-dihydroxybenzylamine as an internal standard. Centrifugation (15,000×g, 30 minutes, 4°C) was performed and the supernatant was passed through a 0.2 µm filter. Twenty microliters of the supernatant were analyzed in the HPLC column (4.6 mm×150 mm C-18 reverse phase column; MC Medical, Tokyo, Japan) by a dual channel coulochem III electrochemical detector (Model 5300; ESA, Inc., Chelmsford, MA, USA) HPLC. Different concentrations of dopamine (DA) standard samples were set to make a standard curve [19].
Determination of serum total antioxidant capacity and serum lipid peroxide or malondialdehyde: Blood samples were obtained from the retro-orbital venous plexus of animals, allowed to clot, centrifuged at 3,000 g for 15 minutes, and the clear, non-hemolyzed supernatant sera were separated for assessment of total antioxidant capacity (TAC) and malondialdehyde (MDA). TAC and MDA were estimated colorimetrically via commercially available kits (Bio-Diagnostics, Giza, Egypt) following the instructions of the kits. Briefly, the hydroxyl radical is formed by the Fenton reaction and via the reaction with the colorless substrate O-dianisidine, it forms the bright yellowish-brown dianisyl radical. On addition of a serum, the antioxidant of the serum depressed the oxidative reactions induced by the hydroxyl radicals and preventing the color change and hence providing an effective determination of TAC [20]. Assessment of MDA based on the reaction of thio-barbituric acid with MDA to produce thio-barbituric acid reactive product. The absorbance of this pink product can be detected at 534 nm [21].
Histological study
Electron microscopy: The rats were deeply anesthetized with thiopental (50 mg/kg intraperitoneally) (IP) [22]. Animals were perfused trans cardially through the left ventricle with Glutaraldehyde. The brain was removed and the midbrain was excised. SN of midbrain was dissected out using a dissecting microscope, fixed in glutaraldehyde cacodylate buffer (PH 7.4) for 24 hours and post fixed in 1% osmium tetroxide for 2 hours and then washed in 0.1 mol/l phosphate buffer (pH 7.4, 15 minutes), and thereafter dehydrated through graded ethanol solutions. The dehydrated blocks were embedded in ethoxy-line resin.
Semithin sections (0.5–1 μm) were cut on the ultra-tome and stained with (1%) toluidine blue (PH 7.3) for light microscopic examination. Ultrathin sections (50–80 nm) were cut from the selected areas in semithin sections, placed on copper grids, double stained with uranyl acetate and lead citrate [23]. The grids were examined in Assiut University-Electron Microscope Unit, using a transmission electron microscope JEOL (JEM-100 CXII, Tokyo, Japan) and photographed at 80 KV.
Morphometric study: Three non-overlapped fields in each of five semi-thin sections of each of 3 rats from each group were used for measuring the following:
1- The number of light cells was counted/field in each section ×1,000
2- The number of dark cells was counted/field in each section ×1,000
3- The surface area of nerve cells area/field µm2.
All morphometric studies were done by using image J which is a Java-based open-source image processing package.
Statistical analysis: The morphometric data of each group were expressed as mean±standard deviation and statistically analyzed using IBM SPSS Statistics for Windows, Version 20.0 (IBM Co., Armonk, NY, USA). The independent t-test was used for comparing the animal groups. A P-values of less than 0.05 were considered significant [24].
Immunofluorescence staining: All animals were perfused transcardially with 50 ml of saline (0.9%), followed by 200 ml ice-cold paraformaldehyde (4%) in 0.1 M, phosphate buffer. The brain was then removed and maintained in the fixative for 24 hours at 4°C. After an overnight incubation in PBS containing 30% sucrose at 4°C, the brain was frozen and then sectioned. Briefly, serial coronal sections of 30 μm thickness were cut using a freezing microtome (Leica CM 1800; Leica Microsystems, Bannockburn, IL, USA) on coated slides. Sections were washed in phosphate-buffered saline (PBS) before application of the primary antibody and also between each immunohistochemistry step [23].
The slices were rinsed with PBS for 5 minutes, permeabilized with PBS-0.1% Triton for 20 minutes, and blocked with PBS containing 3% bovine serum albumin (Sigma) and 0.3%Triton X-100 at room temperature for 90 minutes and labeled with the primary antibody for 3 hours at room temperature. Secondary antibody was used diluted in PBS containing 0.2% BSA for 90 minutes in room temperature. The sections were cover-slipped with mounting media.
The primary antibodies were Rabbit polyclonal anti-tyrosine hydroxylase (1:1,000; Thermo Fisher Scientific, Rockford, IL, USA). Rabbit polyclonal α-Synuclein antibody (1:1,000; Thermo Fisher Scientific) and polyclonal anti-caspase-3 antibody (1:100; Chongqing Biopses, 400020, Chongqing, China) were also used. Caspase 3 is formed of a 32 kDa and zymogen that is cleaved into 17 kD and 12 kDa subunits. The immunogen corresponds to the region of the human caspase 3 p 17 subunit. Caspase 3 plays essesntial roles in programed cell death (apoptosis) where it is responsible for DNA fragmentation and chromatin condensation [25].
The secondary antibodies (Goat anti-Rabbit IgG [H+L], Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 555 [1:300; Thermo Fisher Scientific], Goat anti-Mouse Ig G [H+L], Cross-Adsorbed Secondary Antibody, Alexa Fluor 488 [1:300; Thermo Fisher Scientific]).
Fluorescence images were obtained with Olympus fluorescence microscope equipped with fluorescein and rhodamine filters (BX 51 2007 Tokyo, Japan).
Results
Bioanlysis
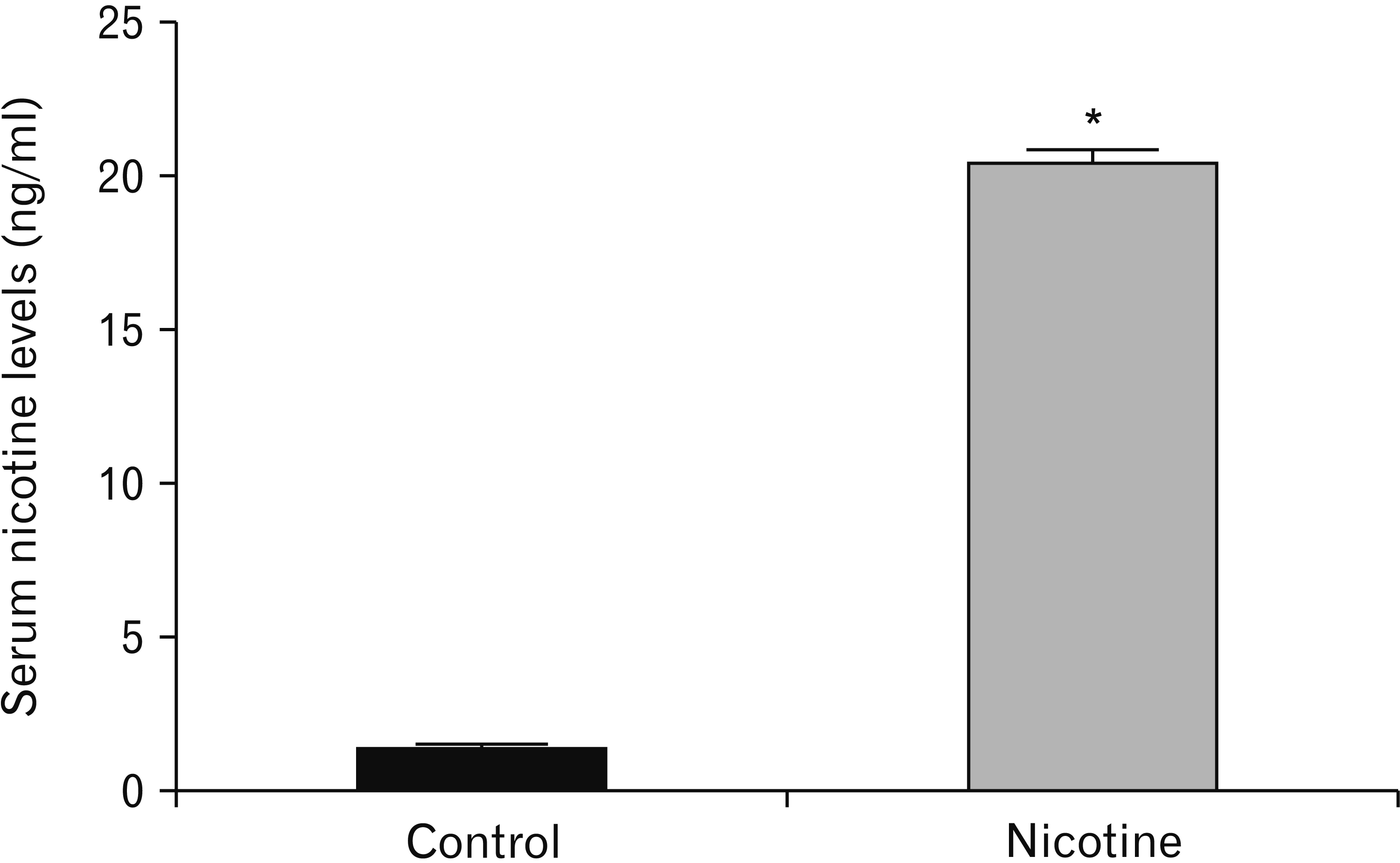
A significant increased level of nicotine was revealed in the serum of animals in the nicotine group when compared with animals in the control group (P<0.0001) (Table 1, Fig. 1).
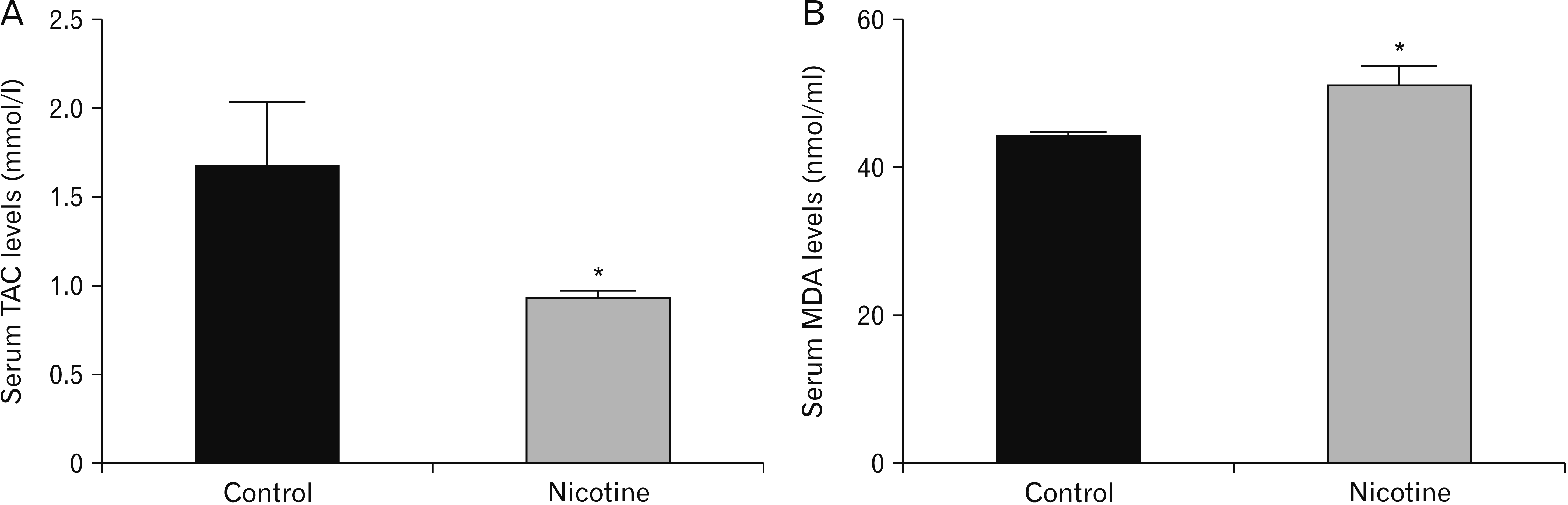
A reduction in the striatal dopamine level in plasma was found in nicotine group compared with the control group, however the difference was statistically insignificant (P>0.05) (Table 2). On the other hand, a significant decrease in the level of TAC was observed in the nicotine group when compared with the control group (P<0.01). In addition, a significant increase in the level of MDA was detected in nicotine group compared with the control group (P<0.01) (Fig. 2).
Morphometric results
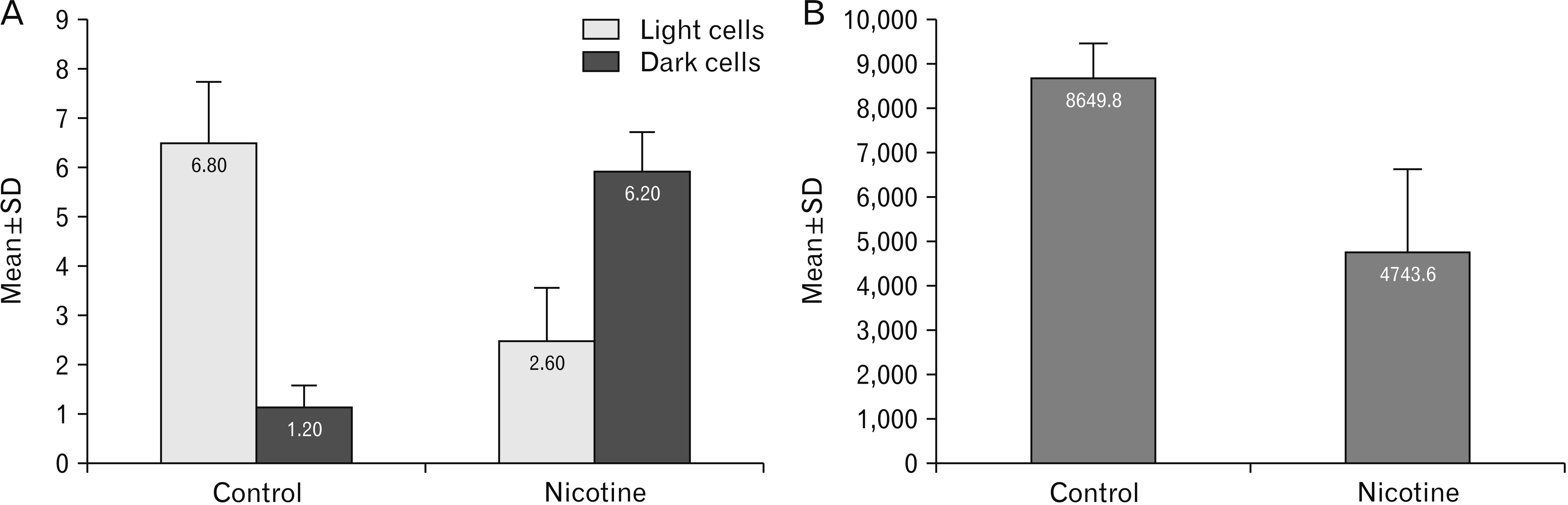
Nicotine treated group revealed a significant reduction in the mean number of light cells per field compared to control group (P-value<0.001). In addition, a significant increase of dark cells was detected in nicotine treated group compared to control (P-value<0.001) (Fig. 3A). The mean surface area of nerve cells/field was significantly reduced in nicotine treated group compared to control (P-value<0.003) (Fig. 3B).
Histological results
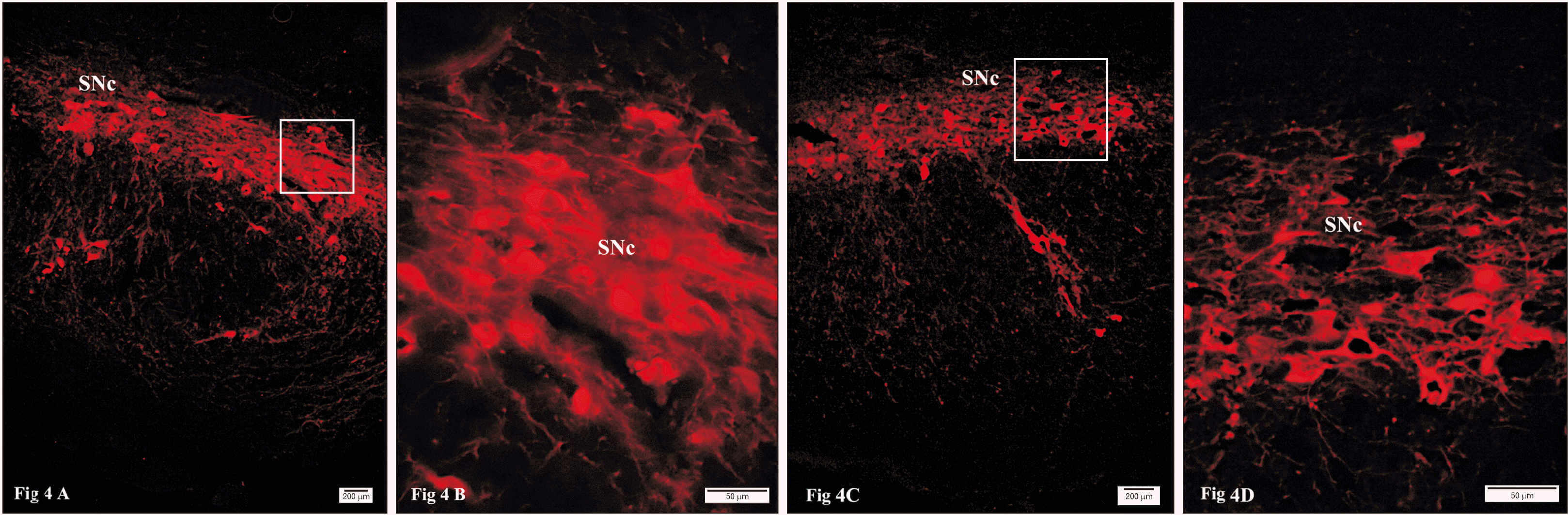
Immunofluorescent staining of control SNc revealed a strong positive reactivity to TH-antibody with numerous TH-positive neurons (Fig. 4A, B). However, this reactivity was less in nicotine-treated group than that detected in control group (Fig. 4C, D). Immunofluorescence staining of caspase 3 was negative in control group (Fig. 5A, B), whereas in nicotine treated group the reaction was positive (Fig. 5C, D). Immunofluorescence for caspase α-synuclein antibodies was negative in control group (Fig. 6A, B), whereas in nicotine treated group the reaction was positive (Fig. 6C, D).
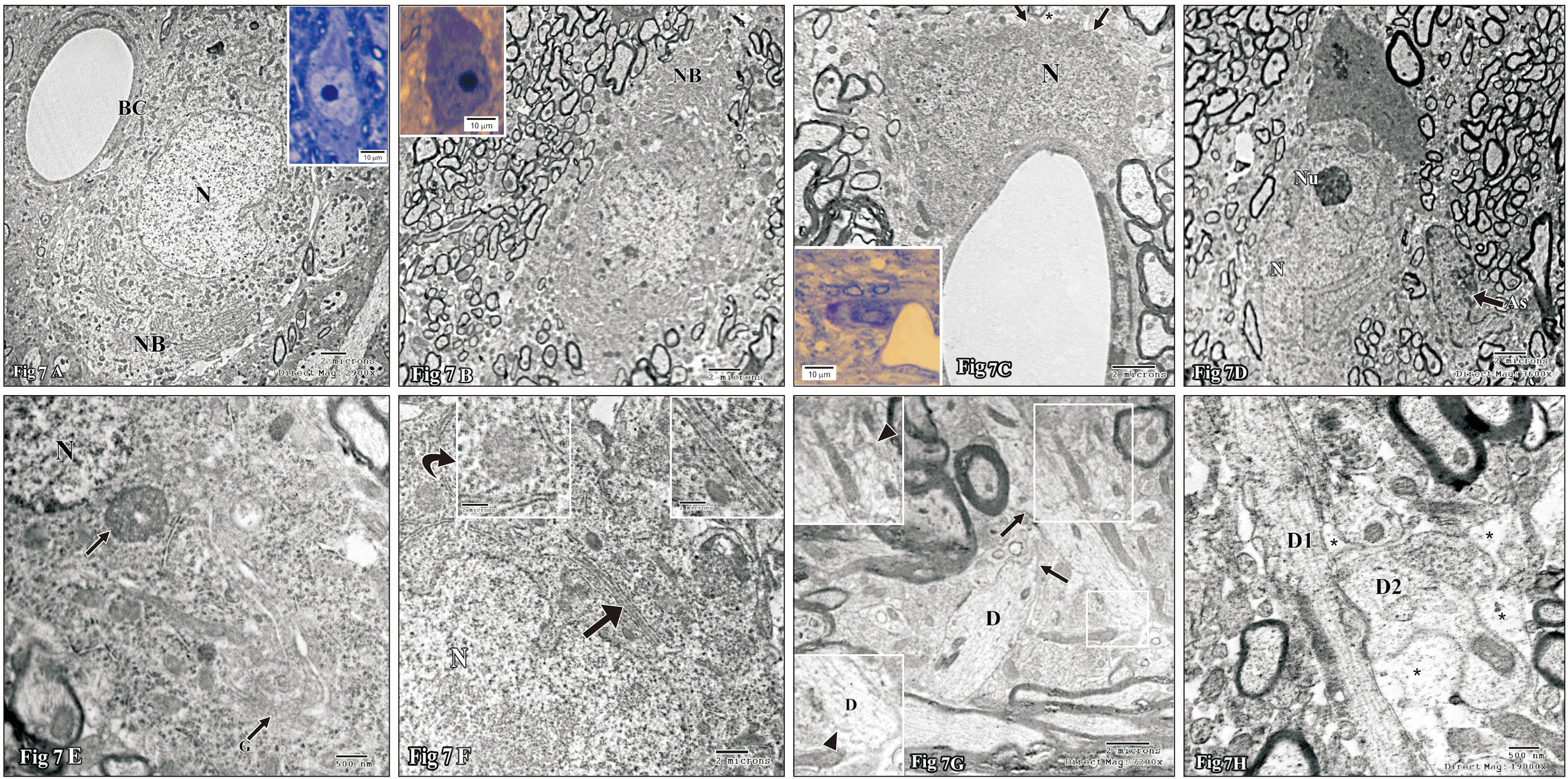
In light microscopy, the compacta neurons of control group were closely arranged in the form of a band of flattened cells distal to SNr. They were lightly stained and variable in shapes (spindle, stellate or fusiform), with vesicular nuclei and prominent nucleoli (Fig. 7A). The surrounding neuropil contained numerous blood vessels, glial cells and nerve fibers. In ultrastructure the neurons revealed euchromatic round to oval central nuclei with prominent nucleoli. The surrounding voluminous cytoplasm contained numerous strands of rough endoplasmic reticulum (RER), mitochondria, well developed Golgi bodies and a few small lysosomes (Fig. 7B). The nerve fibers of the control group exhibited regular outlining and contained mitochondria, microtubules, and neurofilaments (Fig. 7A).
In contrary to the control group, the compacta neurons of nicotine treated group were widely separated from each other and most of them were darkly stained with dark nuclei. However, the nucleoli were still distinct within the nuclei and the cytoplasm was metachromatic (Fig. 7A). Ultrastructure revealed many neurons in variable degrees of electron-density of both the nuclei and cytoplasm with highly prominent nucleoli formed of pars granulosa. Many electron dense neurons revealed a marked increase in cytoplasmic Nissl bodies (Fig. 7B), irregular outlining (blebs) and surrounded by dilated astrocytic processes (Fig. 7C). Other neurons displayed markedly irregular nuclei, fragmented Golgi and non-membrane bound frequently-detectable inclusions (Fig. 7D–F). On high magnification most of them contained fibril-like structures (Fig. 7E, F). Occasionally RER cisternae were dilated and contained secretory material (Fig. 7F), which closely resembled in shape and electron density, the cytoplasmic inclusions (Fig. 7E, F). Many neuronal processes revealed irregular plasma membrane that formed either corrugated or stepwise surface and variable shaped spine projections (Fig. 7G), disorganized microtubules (non-parallel arrangements or widened intervals) and/ or disrupted microtubules (Fig. 7H). The neuropil revealed numerous dendro-dendritic synapses (Fig. 7H).
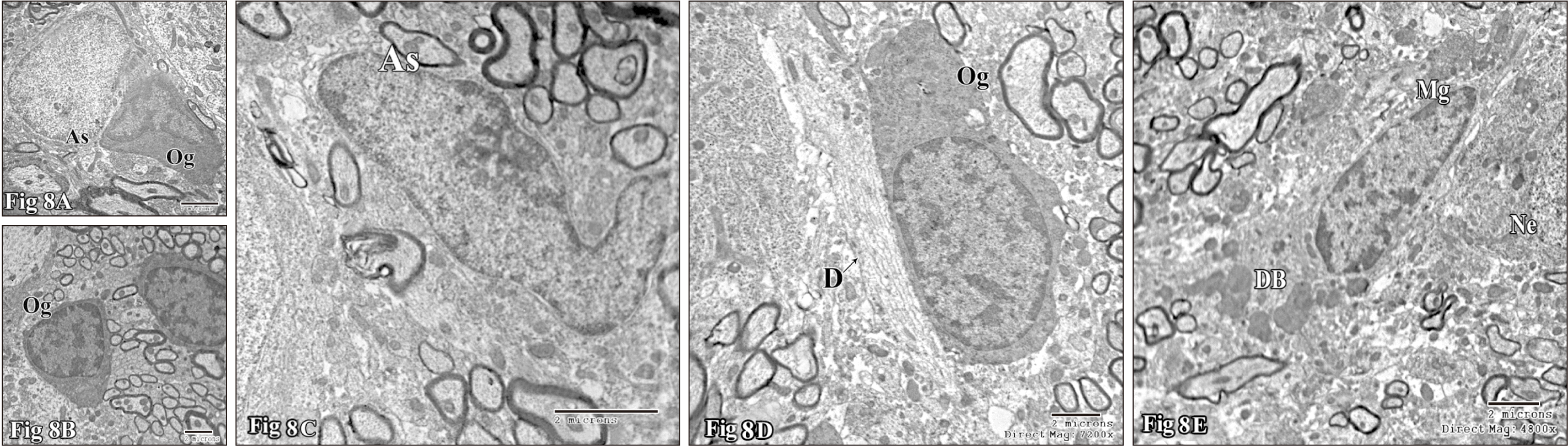
Neuroglial cells in control group included astrocytes, microglia, and oligodendrocytes. The astrocytes exhibited pale nuclei that were usually regular in shape containing peripheral and some scattered heterochromatin. Their cytoplasm was also pale, scanty and contained a few organelles and extended narrow long processes (Fig. 8A). Microglia cells were found either in relation to blood capillaries (Fig. 8A) or within the tissue parenchyma. They exhibited small nuclei with large heterochromatin, surrounded by a scanty cytoplasm. Those present contiguous to neurons, their somata were always separated from those of the neurons by astrocytic processes. Regarding oligodendrocyte, two classes may be described within this group of cells; moderately electron-dense oligodendrocytes (Fig. 8A), which are large cells with moderately electron dense nucleus and cytoplasm, containing well developed organelles, and giving off a fair number of fine processes; and electron-dense oligodendrocytes, with very dense nucleus and cytoplasm, containing a prominent Golgi bodies (Fig. 8B).
In nicotine-treated group the astrocytes were reactive, hypertrophied and abnormally shaped nuclei surrounded by a scanty cytoplasm were occasionally observed (Fig. 8C). Astrocytic end feet processes around the blood capillaries might become swollen (Fig. 9D). Oligodendrocytes might be enlarged with numerous organelles and/or numerous dense inclusions and rod elongated nuclei (Fig. 8D) and usually encountered as two identical adjacent cells. Reactive microglia characteristically became hypertrophied with relatively electron-dense cytoplasm containing numerous electron dense bodies and elongated nuclei (Fig. 8E).
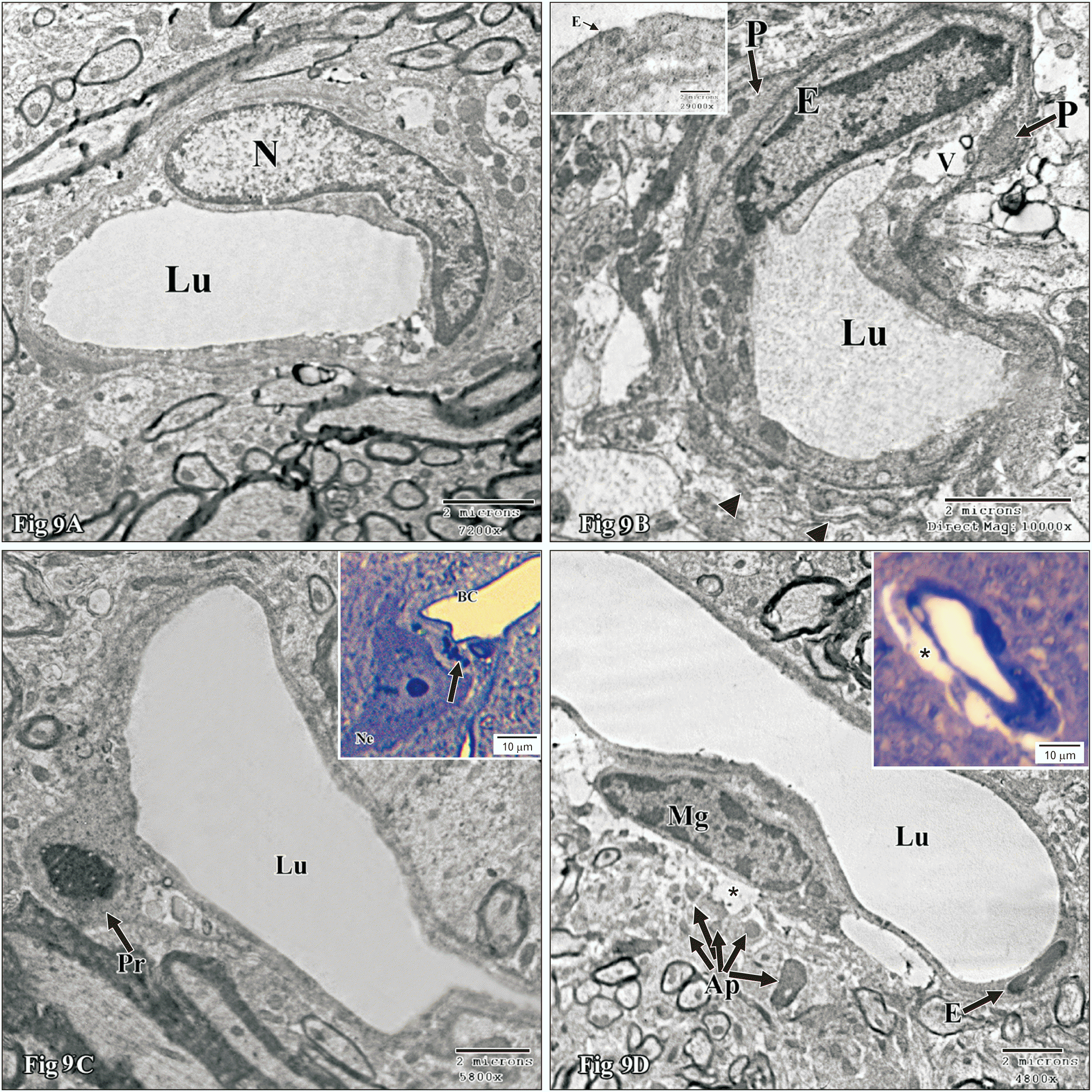
The blood capillaries in the control group were lined by highly flattened endothelial cells with flattened nuclei and a few organelles (Fig. 5A). Endothelial cells were connected by tight junctions and enveloped by pericytes. In nicotine treated group, the blood capillaries might show irregular lumen, attenuated endothelial cells and/or apoptotic pericytes (Fig. 9A). The blood capillaries might reveal an abnormally large irregular (Fig. 9A), attenuated or apoptotic endothelial lining and irregular lumina (Fig. 9B, C). In addition, the vascular endothelium might exhibit indistinct junctional complex or fenestrae (Fig. 9B). The surrounding pericytic (Fig. 9C) and/or perivascular cells might also reveal apoptosis (Fig. 9D).
Discussion
This work revealed that nicotine use (11 days) might produce injurious effects on both histological structure and function of the SNc. This would influence the striatum function as DA neurons located in the SNc primarily project to form the so-called nigrostriatal pathway, which predominantly regulates motor function. In support with our suggestion, we found that nicotine use produced no increase in DA level (the increment was insignificant) and a decrease in immune reactivity for TH, the marker for dopamine neurons. The literature also revealed that greater lifetime use of cigarettes (measured in pack-years) was associated with smaller left nucleus accumbens area volume (P=0.018) and larger left putamen volume (P=0.025) [13]. The presence of numerous electron-dense neurons revealing presence of multiple Nissle bodies along with prominent nucleoli (formed mainly of pars granulosa) indicate neuronal excitation. It is declared that nicotine exerts an excitatory action in cell body of neurons at the level of nigral DA nerve cells [26]. So, we propose that nicotine use in control animals might induce stimulation followed by exhaustive degeneration.
Reduced dopaminergic neurons, that we observed in nicotine treated group, occurred most probably by apoptosis. This is confirmed by immunohistochemistry using anti caspase-3 antibodies and by ultrastructure. The apoptotic features included an increase in neuronal electron density, their shrinkage and blebs of the neurons outlining. In addition, some cells exhibited breakdown of the Golgi apparatus which is considered an early event during apoptosis [27] as the Golgi localized caspase-2 and caspase-3 mediate cleavage of several Golgi-associated proteins That early event coincided with release of cytochrome c from mitochondria. Significantly, Golgi fragmentation preceded major changes in the organization of both the actin cytoskeleton and microtubules [28].
Coinciding with our findings numerous investigations carried on neuronal interactions of nicotine have highlighted the adverse potentials of this agent on some brain regions. In rat temporal cortices nicotine induced neuronal apoptosis [2]. In the prefrontal cortex, nicotine administration decreased the size of perikria and number of dendritic spines [29]. In addition; loss of cell layers thickness and their cellular constituents was induced in the cerebellum by nicotine [30]. Hippocampal and striatal neurodegenerative changes have been recorded as well under chronic nicotine administration [3]. In auditory cortex, nicotine produced an injurious dose related effect on the neurons [31].
Neuronal cells are specifically vulnerable to damage by peroxidation owing to their high oxygen utilization, high content of oxidizable polyunsaturated fatty acids, and the existence of redox-active metals (Cu, Fe) [32]. Decreased TAC detected in this work implies inability to inactivate the ROS. Along with the increase in MDA, the lipid peroxidation marker, we can suggest that neuronal activation for long period can result in their damage driven by biochemical alterations. In support of this suggestion, nicotine is found to block anti-oxidant enzymes, thereby increasing peroxidation of lipids, and formation of oxygen free radicals. Free radicals in the tissues are able to trigger cytoplasmic membrane damage and DNA fragmentation. An association between excessive formation of oxygen free radicals and the development of diseases such as tumors, hyperglycemia, arteriosclerosis, stroke, and neurodegenerative disorders has been reported. According to Cobley et al. [32], the rat temporal cortices revealed reduced antioxidant enzymes activity, and increased lipid peroxidation and oxygen free radicals. Moreover, plasma TAC level decreased in bidi workers as per exposure to nicotine [33], and the ROS levels increased in brain regions concerned with reward after recurrent exposure to a variety of misused drugs, including nicotine [34]. In addition, it has been reported that nicotine (2.5 mg/kg), reduced the total serum antioxidant capacity level with growth in the rate of ROS from nicotine at pancreases strictures of mice [35].
The positive reaction for antibodies against α-synuclein observed in this study in nicotine group could be correlated with the presence of cytoplasmic assembles (aggregates) detected by electron microscopy and the secretory material detected inside dilated RER. So, we propose that chronic nicotine use might induce cytoplasmic aggregations and formation of inclusion bodies. Cytoplasmic inclusions more or less similar to those found here are formed following oxidative insult [36]. In their study, they tested the possibility that exposure of cells to patho-physiologically reasonable fluxes of reactive oxygen may induce the formation of protein aggregates in HEK 293 cells transfected with wild-type or mutant human α-synuclein [36]. In further support of our suggestion neurons of high-expresser human α-synuclein mice (2–3 months of age) showed electron-dense deposits (0.1–0.25 mm in diameter) associated with the RER [37]. Moreover, Miraglia et al. [38], has found that α-Synuclein associates with specific membranes in neurons such as that of synaptic vesicles and some cellular organelles, like the RER/Golgi and the mitochondria. Since nicotine induced a decrease in TAC level, the aggregations of α-synuclein could be attributed to the cellular adversities resulting from oxidative stress.
Under optimal conditions, α-Synuclein functions in the normal brain as a soluble monomer. These are thought to mediate its physiological function in presynaptic terminals [39]. However, the forms of α-synuclein oligomers and protofibrils are considered highly toxic. As they accumulate at presynaptic endings, they affect many steps of neurotransmitter release.
The dendro-dendritic reactive synaptogenesis detected in nicotine group, most probably, function to compensate for and/or restore partial functionality to the affected region as suggested by [40]. Those investigators proposed that this type of dendritic reactive synaptogenesis is thought to occur so as to re-saturate the region which has become vacant post-synaptic site following neurodegeneration caused by deafferentation or axotomy so that a partial functionality is restored [41].
Dilated astrocytic end-feet which surround the blood capillaries could probably be secondary to disrupted blood–brain barrier. In support of this suggestion, Huang and his coworkers [12] reported that the blood circulating brain microvascular endothelial cells (BMECs, an indicator of vascular damage), increased in mice after nicotine treatment [42]. So, dysfunction of BMECs, which are a major component of the BBB, could be caused by nicotine.
Ischemia that might arise secondary to reduced blood flow could also account for the dilated astrocytic processes. This suggestion coincides with Petito and Babiak [43], work who reported that the most punctual reaction to anoxia-ischemia is conspicuous swelling of astrocytes which happens at first in end-feet around vessels. Since nicotine might induce vasoconstriction and consequently a decrease in cerebral blood flow (CBF) [44]. Loss of pericytes could result either directly from oxidative stress or indirectly through reduced cerebrovascular flow by nicotine. It is evident that pericytes are particularly sensitive to damage during pathological insults such as ischemia. According to Hall et al. [45], rat brain ischemia would induce persistent pericytes contraction and subsequent pericytes death. Pericytes have significant functions in BBB formation and maintenance, monitoring capillary diameter and CBF adjustment [46].
In conclusion, chronic nicotine administration in a dose (1.5 mg/kg body weight) might induce a negative influence on SNc. The reduced TAC, neuronal apoptosis and vascular adverse effects most probably were secondary to an oxidative stress.


 XML Download
XML Download