

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Moraxella catarrhalis, a gram-negative aerobic diplococcus, has been recognized as an increasingly important pathogen in respiratory infections [1-3]. M. catarrhalis usually causes sinusitis and otitis media in children and lower respiratory tract infections and exacerbation of chronic obstructive pulmonary disease in adults [4]. Currently, penicillin is most commonly used to treat M. catarrhalis infections [5]. However, M. catarrhalis isolates are frequently resistant to ampicillin owing to the production of β-lactamase (typically BRO-1 or BRO-2) [3]. The positive rate of β-lactamase in M. catarrhalis is > 90% in many countries [6, 7]. This high prevalence of β-lactamase production in M. catarrhalis isolates led to the initial use of tetracyclines as an alternative to β-lactamase antimicrobials for the treatment of respiratory illnesses caused by M. cattarhalis [8]. However, the tetracycline resistance rate among M. catarrhalis isolates fluctuates between 0–30% [9].
Tetracycline resistance is usually attributed to the acquisition of mobile genetic elements carrying tetracycline-specific resistance genes (encoding an efflux pump, a ribosome protective protein, or a drug inactivating enzyme), mutations within the ribosomal binding sites, and/or chromosomal mutations leading to an increased expression of intrinsic resistance mechanisms [10]. Since Mendez, et al. [11] first studied the genetic mechanism of plasmid-mediated tetracycline resistance genes in Enterobacteriaceae and Pseudomonas in 1980, various tetracycline resistance genes have been discovered in gram-positive and gram-negative bacteria [12]. These genes often combine with mobile components, such as transferable plasmids, transposons, conjugative transposons, and integrants, resulting in the extensive spread of resistance genes among flora [12, 13]. Some studies on tetracycline resistance mechanisms in M. catarrhalis have identified tetM, tetL, tetW, tetO, and tetQ as the genes involved in tetracycline resistance in Neisseria spp., which show high similarity with M. catarrhalis with respect to phenotypic characteristics and ecological niche. Reports suggest that tetM, tetL, tetW, tetO, and tetQ could be involved in tetracycline resistance in M. catarrhalis, while another study has shown that the tetB is also involved in tetracycline resistance [12–14].
Owing to the severe problem of resistance to traditional tetracyclines, such as doxycycline and minocycline, new tetracyclines, such as tigecycline, eravacycline, and omadacycline, have been developed recently and have shown excellent activity against gram-positive and gram-negative bacteria [15, 16]. However, the susceptibility of different microbes to these new tetracyclines and their mechanism of action against M. catarrhalis have not been well elucidated. Tigecycline resistance has been linked to genetic mutations affecting the 30S ribosomal subunit of the tetracycline binding site, including mutations affecting the genes encoding 16S rRNA (four copies) and ribosomal protein S10 [17]. Moreover, these gene mutations have not been shown to increase the minimum inhibitory concentrations (MICs) of omadacycline and eravacycline against M. catarrhalis. Thus, this study explored the susceptibility of M. catarrhalis to these novel tetracyclines and investigated the possible resistance mechanisms.
MATERIALS AND METHODS
Bacterial strains
A total of 207 non-duplicate M. catarrhalis isolates were collected from different inpatients at Shenzhen Nanshan People’s Hospital Shenzhen University, China, between January 1st, 2012 and December 31st, 2017 from the following sources: 161 from sputum (77.8%), 43 from throat swabs (20.8%), two (0.9%) from blood, and one (0.5%) from bronchoalveolar lavage fluid. The isolates were identified using the Phoenix 100 automated microbiology system (Becton Dickinson [BD], Franklin Lakes, NJ, USA), and their identities were confirmed by matrix-assisted laser desorption ionization time-of-flight mass spectrometry (IVD MALDI Biotyper; Bruker, Bremen, Germany). Staphylococcus aureus ATCC 29213 was used as the quality control strain.
All procedures involving human patients in this retrospective study were approved by the ethics committee of Shenzhen Nanshan People’s Hospital, according to the ethical standards of Shenzhen University and the 1964 Helsinki declaration and its later amendments, or comparable ethical standards (approval number: SZNS2017015). For this type of study, formal consent was not required.
Chemicals and antimicrobial susceptibility testing
Doxycycline (catalog No. HY-N0565), minocycline hydrochloride (catalog No. HY-17412), tigecycline hydrochloride (catalog
No. HY-B0117A), eravacycline (catalog No. HY-16980), and omadacycline (catalog No. HY-14865) were purchased from MedChemExpress (MCE, Shanghai, China).
Antimicrobial MICs were determined using the broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI) document M45-A2 [18]. Overnight bacterial cultures were diluted 1:100 in 2 mL of 1 × Mueller-Hinton broth (MHB). Following 3 hours of incubation at 37°C, 10.82 ×g, the colony suspension (corresponding to a 0.5 McFarland standard, 1.0–1.5 × 108 colony forming units [cfu]/mL) was diluted 1:100 in 1 mL 2 × MHB. Aliquots (100 μL) of the colony suspension (1.0–1.5 × 106 cfu/mL) were inoculated into polystyrene microtiter plates (Costar3599; Corning, NY, USA) containing 100-μL aliquots of tigecycline, eravacycline, or omadacycline (0.0078, 0.0156, 0.031, 0.06, 0.125, 0.25, 0.5, 1, 2, 4, or 8 mg/L). Following 24 hours incubation at 35°C, the MICs were calculated as the drug concentration in the well without obvious bacterial precipitation. At present, there are no unified criteria for determining the antimicrobial susceptibility of M. catarrhalis. Thus, we used the European Committee on Antimicrobial Susceptibility Testing (EUCAST)-recommended MIC breakpoints for M. catarrhalis. Isolates with an MIC ≤ 1 or > 2 mg/L were considered susceptible or resistant, respectively, to doxycycline or minocycline.
In vitro induction of tigecycline-, eravacycline-, or omadacycline-resistant isolates
To explore the possible resistance mechanisms of M. catarrhalis against the new tetracyclines, tigecycline-, eravacycline-, or omadacycline-resistant isolates were induced under in vitro selection pressure. Three M. catarrhalis isolates (MC1 [tetB-, tetL-], MC2 [tetB-, tetL+], and MC3 [tetB+, tetL-]; the genotypes were determined by PCR detection of tetracycline-specific resistance genes) with low tigecycline (0.125 mg/L), eravacycline (0.125 mg/L), and omadacycline (0.25 mg/L) MICs were used to induce the resistant isolates. The parental isolates were subcultured serially in MHB containing gradually increasing concentrations of tigecycline, eravacycline, or omadacycline. The initial inducing concentration was 0.5 × the initial MIC, which was then successively increased to 1 ×, 2 ×, 4 ×, 8 ×, 16 ×, 32 ×, or 64 × the initial MIC [19]. Isolates were cultured for 3–5 passages before being transferred into the next concentration. Isolates from the passages of each concentration were stored at -80°C in MHB containing 40% glycerol to identify genetic mutations and to test subsequent MIC.
PCR detection of the tet genes and mutations in the 16S rRNA and the 30S ribosomal protein S10 genes after 160 days of induction
DNA was extracted and purified from all isolates using the DNeasy Blood and Tissue Kit (Qiagen, Shanghai, China), according to the manufacturer’s protocol for gram-negative bacteria. tet primers described by Collins, et al. [20] and Villedieu, et al. [21] were used. The primers used for the 16S rRNA and 30S ribosomal protein S10 genes are listed in Table 1. Mutations in the 16S rRNA and the 30S ribosomal protein S10 genes were detected by PCR amplification and sequence alignment [17]. PCR amplification was performed in a total volume of 50 μL, containing 2 × PCR Master Mix (TIANGEN Biotech Beijing Co., Ltd., Beijing, China), 0.5 μmol/L of each primer, and 1 μL of template DNA. The cycling conditions were as follows: 94°C for 5 minutes; 30 cycles of 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 30 seconds; and a final 10-minute extension step at 72°C. Each PCR set included a no-template control and a positive control. The amplification products were analyzed by electrophoresis on 1.0% agarose gels.
RESULTS
Susceptibility of M. catarrhalis to tetracyclines
The MIC distributions of the tetracyclines among the M. catarrhalis isolates are summarized in Table 2; eight isolates had a doxycycline MIC ≥ 1 mg/L, and four isolates had a minocycline MIC ≥ 1 mg/L. Applying the EUCAST interpretive criteria, 97% of the isolates were susceptible to doxycycline and 98% were susceptible to minocycline.
Of the three new tetracyclines, eravacycline had a lower MIC50 (0.06 mg/L) than tigecycline (0.125 mg/L) or omadacycline (0.125 mg/L) and MIC90 (0.125 mg/L) for the 207 M. catarrhalis isolates (Table 2). The number of isolates with an MIC of 0.06 mg/L was the highest for eravacycline (156/207, 75.4%), followed by tigecycline (71/207, 34.3%; P < 0.001), and omadacycline (33/207, 15.9%; P < 0.001). Only two isolates had tigecycline or eravacycline MICs ≥ 1 mg/L, while five isolates had an omadacycline MIC ≥ 1 mg/L.
Tetracycline resistance-related tet genes in M. catarrhalis
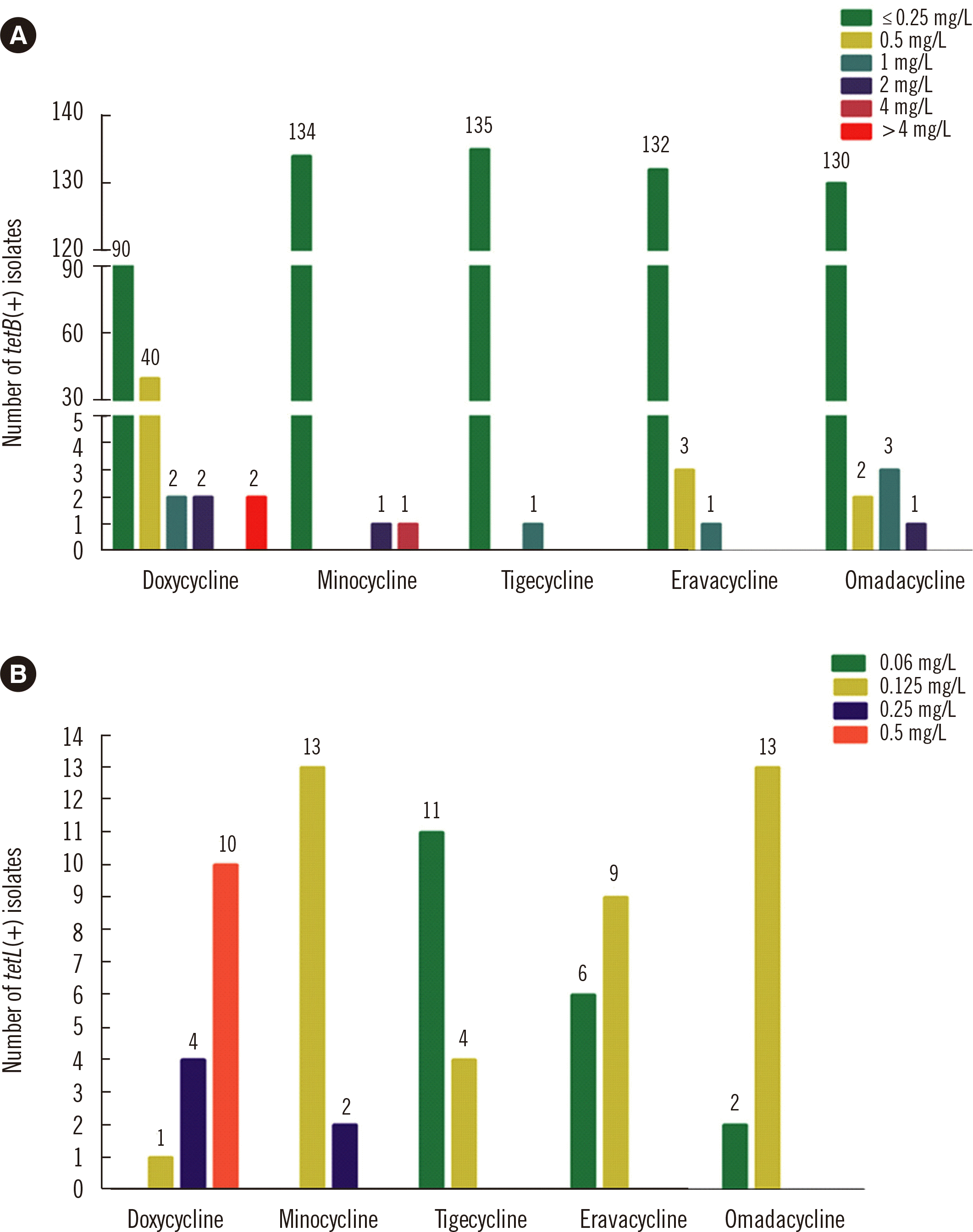
We found that 136 (65.7%) M. catarrhalis isolates harbored tetB, and 15 (7.2%) isolates harbored tetL; tetM, tetW, tetQ, and tetT were not detected in any of the isolates. Only four M. catarrhalis isolates harboring tetB had a high doxycycline MIC ( ≥ 2 mg/L), and two isolates had a high minocycline MIC ( ≥ 2 mg/ L). Similarly, of the tetB-positive isolates, only one had a high tigecycline or eravacycline MIC ( ≥ 1 mg/L), and only four had a high omadacycline MIC ( ≥ 1 mg/L) (Fig. 1A). None of the M. catarrhalis tetL-positive isolates demonstrated high tetracycline MICs ( ≥ 1 mg/L) (Fig. 1B).
Tigecycline-, eravacycline-, and omadacycline-resistant isolates were induced under in vitro selection pressure
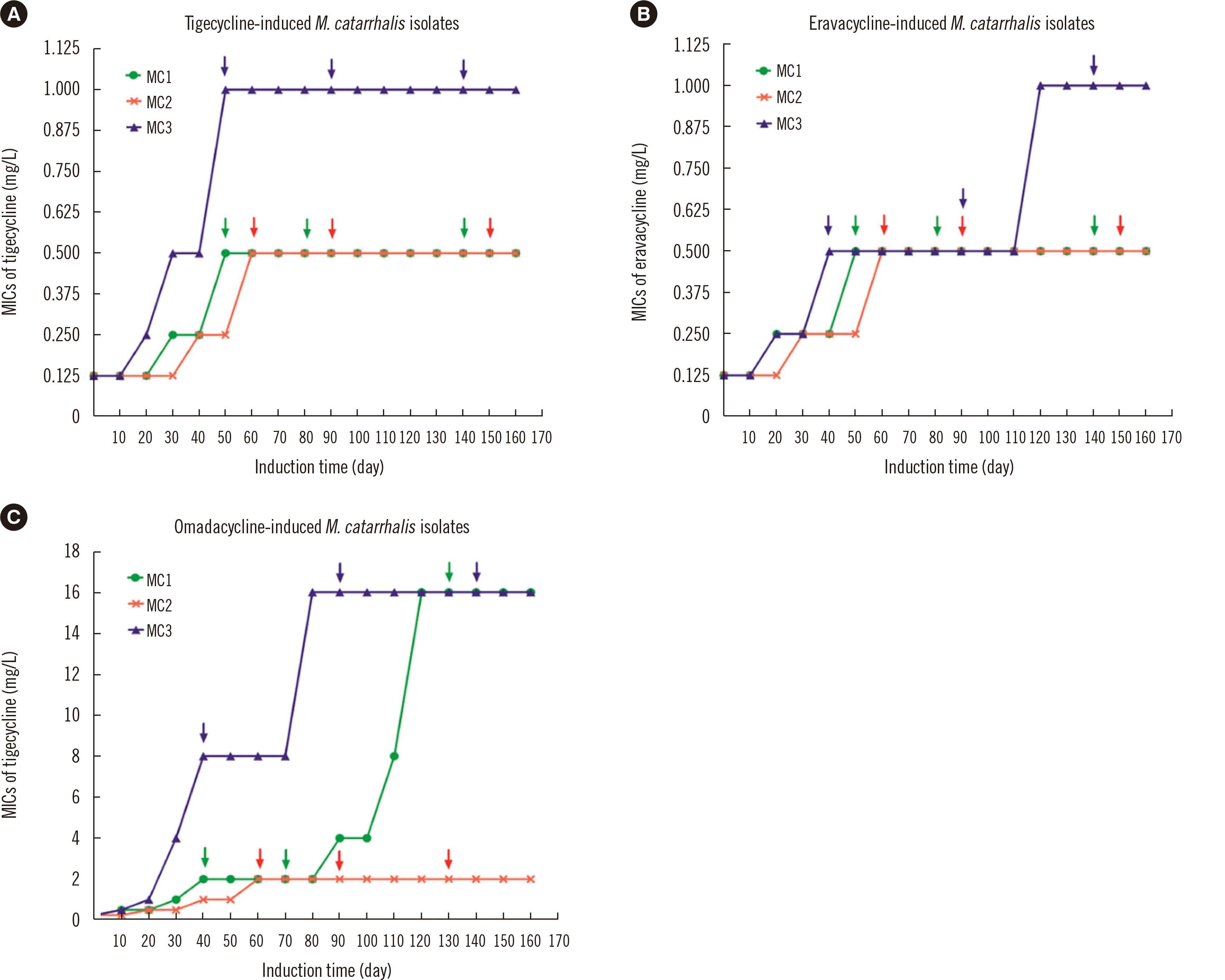
It was difficult to induce resistance to tigecycline or eravacycline in the three parental M. catarrhalis isolates; the MICs of the MC1 and MC2 isolates only increased four-fold compared with the initial MICs, while the MICs of MC3 increased eight-fold compared with the initial MICs (Fig. 2). Compared with tigecycline and eravacycline MICs, the omadacycline MICs of the MC1 and MC3, but not the MC2 isolates, easily increased 64-fold compared with the initial MICs after 160 days of induction.
Detection of 16S rRNA mutations in tigecycline-, eravacycline-, and omadacycline-resistant isolates
Interestingly, the tigecycline-induced resistant isolates showed decreased susceptibilities to the other two new tetracyclines, eravacycline and omadacycline. A similar effect was observed in the eravacycline-induced resistant isolates (Table 3). However, no mutations in the 16S rRNA genes were identified in these tigecyclineor eravacycline-induced resistant isolates. The omadacycline-induced resistant isolates also demonstrated significantly reduced susceptibilities to tigecycline and eravacycline, and novel mutations in the 16S rRNA genes (C1036T and/or G460A) were identified. Among these omadacycline-induced resistant isolates, increased RR copy number of the 16S rRNA genes with mutations was associated with increased resistance to omadacycline. No S10 protein mutations were detected in the new tetracycline-resistant isolates (data not shown).
DISCUSSION
In the present study, > 97% of the M. catarrhalis isolates were susceptible to doxycycline and minocycline and also had low MICs for the three new tetracyclines (tigecycline, eravacycline, or omadacycline). These results support the potential application of the novel tetracyclines in the treatment of M. catarrhalis infections.
The MIC50/90 of omadacycline (0.125/0.125 mg/L) in this study was similar to those reported previously [22]. However, the tigecycline MIC50 (0.125 mg/L) of the 207 M. catarrhalis isolates in this study was higher than the previously reported data (0.004 mg/L) [23]. This may be due to environmental and regional variation. Although a recent study has demonstrated that Klebsiella pneumoniae isolates from China have higher eravacycline MICs than isolates from Europe and the United States, the eravacycline MIC for M. catarrhalis isolates in our study was low (MIC50 at 0.06 mg/L) and lower than that of tigecycline or omadacycline [24].
The mechanisms of tetracycline resistance in gram-positive and gram-negative pathogens include ribosomal protection proteins and efflux pumps [10]. Overexpression of the ribosomal protection protein gene tetM and the efflux pump gene tetL has been reported to reduce the susceptibility of Enterococcus faecium to tigecycline [25]. However, our results showed that the efflux pump genes tetB and tetL did not affect the susceptibility of the M. catarrhalis isolates to tigecycline, eravacycline, or omadacycline. This suggests that either efflux pumps are not the main tetracycline resistance mechanism in M. catarrhalis or that the three new tetracyclines can overcome tetB-mediated resistance in M. catarrhalis.
Crystallographic studies of the 30S ribosomal subunit of Thermus thermophilus revealed the presence of at least six tetracycline binding sites in 16S rRNA [26-28]. Several bacterial species have exhibited higher binding affinities for tigecycline and omadacycline than for tetracycline, and mutations in 16S rRNA that affect the tetracycline binding sites of the 30S ribosomal subunit have been shown to confer tetracycline and tigecycline resistance [28-31]. We investigated mutations in the four copies of the 16S rRNA gene in M. catarrhalis, induced under in vitro tigecycline, eravacycline, or omadacycline pressure. Consistent with previous studies, we found that M. catarrhalis isolates with higher copy numbers of 16S rRNA with genetic mutations tended to have greater omadacycline resistance [17]. The mutational characteristics of the 30S ribosomal protein S10 were consistent with previous results demonstrating that mutations occurred at a relatively low rate in isolates with omadacycline-induced resistance [32]. These results indicate that the role of S10 in the emergence of omadacycline resistance needs to be further studied.
Interestingly, it was very difficult to induce tigecyclineor eravacycline-resistant M. catarrhalis isolates in this study. For example, the tigecycline or eravacycline MICs of the M. catarrhalis isolates increased only eight-fold compared with the initial MICs after 120 days of induction and remained eight-fold higher than the initial MICs even after 160 days of induction. In contrast, in S. aureus, the solithromycin (a novel fluoroketolide antimicrobial) MIC could be induced 128-fold compared with the initial MIC by < 30 days of induction [19]. These results indicate that M. catarrhalis cannot easily develop resistance against tigecycline and eravacycline, and this finding is valuable with respect to antimicrobial application against M. catarrhalis. In contrast, M. catarrhalis may acquire omadacycline resistance more readily, as the omadacycline MIC increased 64-fold compared with the initial MIC after 120 days of induction.
This study has potential limitations. All strains were collected from inpatients, and thus, the in vitro activity of the novel tetracyclines against M. catarrhalis in outpatients remains unknown. Further studies are needed to reveal M. catarrhalis susceptibility from outpatients to tigecycline, eravacycline, and omadacycline.
In conclusion, the traditional tetracyclines, doxycycline and minocycline, and the three novel tetracyclines, tigecycline, eravacycline, and omadacycline, exhibited robust antimicrobial effects against clinical M. catarrhalis isolates from China. The present data contributes to understanding potential resistance mechanisms that may impact the clinical application of omadacycline.
 XML Download
XML Download