

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Tinnitus, defined as perception of sound in the absence of corresponding external auditory stimuli, has a reported prevalence of 10%–15% in adults1 and about 20% of adults with tinnitus require clinical intervention.2 The most widely accepted theory regarding the central origin of hyperexcitability in tinnitus is an imbalance between excitation and inhibition. After acute insult, such as acoustic trauma or exposure to ototoxic drugs, the spontaneous activity increases in the auditory nerve, inferior colliculus, and auditory cortex.34 The dorsal cochlear nucleus, inferior colliculus, and primary auditory cortex neurons are hyperexcitable during the chronic phase.56
Brain mechanisms have been suggested to be involved in the development of tinnitus, and a number of animal models have been developed to test these hypotheses by measuring tinnitus objectively. Tinnitus was reported to be induced in rodents treated with sodium salicylate, and was associated with the alterations of neural activity in the central auditory system.7 Additional animal models have been reported, including induction of tinnitus by intense sound exposure.89
A behavioral model based on the acoustic startle reflex test was recently adopted and has been widely used for objective measurement of tinnitus.10 Briefly, acoustic startle can be evoked by noise and the magnitude of this startle can be suppressed by a gap of silence in the background sound that precedes the noise. If tinnitus is present, the sound of tinnitus will fill the gap and reduce the degree to which it suppresses the acoustic startle response.11 The ratio between the magnitude of the startle stimulus presented alone (no-gap trial) and trials in which a gap precedes startle stimulus (gap trials) is calculated as the gap-prepulse inhibition of the acoustic startle (GPIAS) ratio12 and reflects gap detection. Reduced inhibition by gaps embedded in specific background noise frequencies is assumed to reflect tinnitus frequency.13
It was reported that damage to cochlear hair cells was essential for the pathogenesis of tinnitus,14 indicating that cochlear lesions result in elevation of hearing thresholds and initiation of tinnitus. Therefore, the early stage of tinnitus could be controlled by appropriate therapy, such as steroid administration, if the cochlear damage remains reversible. The effects of steroids include immunosuppression, antiinflammation, and sodium reabsorption. Intratympanic steroid administration was reported to be equivalent to high-dose oral prednisone therapy as primary therapy in the treatment of inner ear disorders.15
This study was designed to identify acute tinnitus and to evaluate the efficacy of steroid administration for noise-induced acute tinnitus by measuring the GPIAS value in an animal model.
Go to : 
METHODS
Subjects
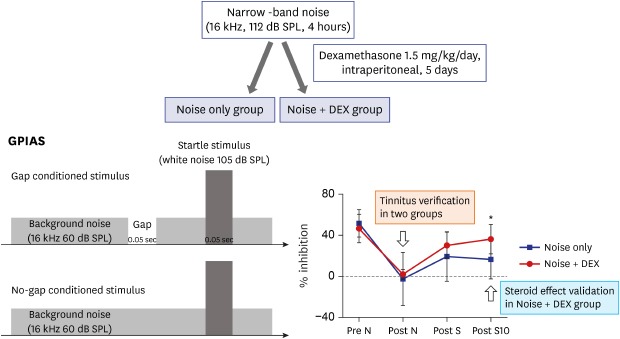
A total of 19 male Sprague-Dawley (SD) rats (8 weeks old, 250–300 g) were used in this study. All SD rats were maintained under conditions of controlled temperature (22°C ± 2°C), humidity (70%–75%), and light/dark cycle (lights on at 8:00, lights off at 20:00). Rats were divided into two groups: the noise group (n = 7) and the noise + dexamethasone (DEX) group (n = 12) (Fig. 1).
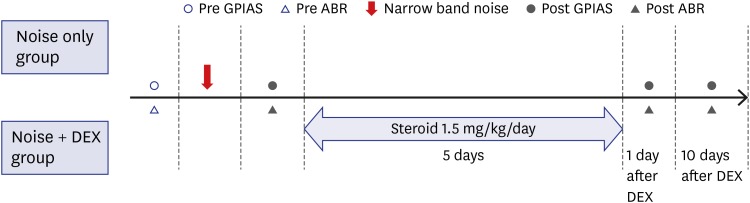 | Fig. 1Experimental schedules. Nineteen rats were divided into the noise group (n = 7), and noise + DEX group (n = 12) injected intraperitoneally with 1.5 mg of DEX/kg daily for 5 days after completion of noise exposure. GPIAS and ABR were recorded just before and after noise exposure. ABR measurement was repeated 1 and 10 days after completion of DEX administration.DEX = dexamethasone, GPIAS = gap-prepulse inhibition of the acoustic startle, ABR = auditory brainstem response.
|
Noise exposure
Rats were exposed for 4 hours to narrow-band noise centered at 16 kHz from a sound generator (Sine Random Generator Type 1027; Brüel & Kjær, Copenhagen, Denmark), amplified (R300 Plus Amplifier; Inter-M, Seoul, Korea) to 112 dB SPL, and played through a speaker (#84234xx; Electro-Voice, Burnsville, MN, USA).
GPIAS
Each rat was placed in an acoustically transparent unconstrained type enclosure in an anechoic chamber (startle reflex chamber). The anechoic chamber had acoustic open cell foam walls and the unconstrained type enclosure rested on an accelerometer, which measured the startle response. All studies were conducted in a soundproof chamber (Fig. 2).
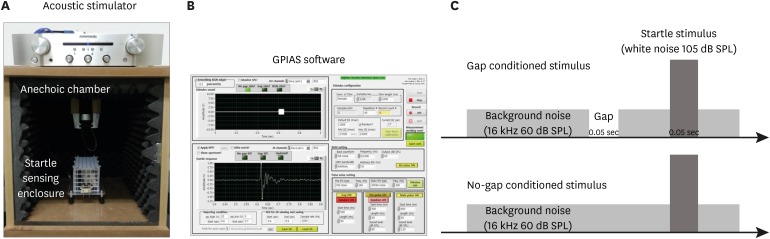 | Fig. 2Components of the GPIAS measurement system and acoustic stimuli used for GPIAS. (A) This system was composed of a soundproof chamber with acoustic stimulator and a startle reflex-sensing enclosure. (B) Implemented PC-based graphical user interface screen for measuring GPIAS values. Schematic view of pulse pattern of (C) gap and no gap.GPIAS = gap-prepulse inhibition of the acoustic startle.
|
GPIAS sessions were composed of 30 gap trials and 30 no-gap trials, which were presented in random pairs. Trials were separated by conditional random interstimulus interval. This system measures the animal startle response only when an animal is in the stable status by sensing the animal's movements using an accelerometer in real-time to minimize motion artifacts. A 2-minute acclimation period was allowed at the beginning of each session. Each gap trial was composed of 60-dB SPL continuous narrow band background noise centered at 16 kHz, and a startle stimulus (single broadband noise burst, 110 dB SPL, 20 ms length) preceded by a long gap of 50 ms ending 100 ms before the onset of the startle stimulus. In the no-gap trials, the background sound was continuous with no silent period preceding the startle stimulus. GPIAS was measured in all rats just prior to and 1 day after noise exposure and 1 and 10 days after completing steroid administration.
Systemic steroid administration
The noise + DEX group received intraperitoneal steroid administration daily for 5 days (1.5 mg/kg/day) after completing noise exposure.
Auditory brainstem response (ABR)
ABR was recorded in all rats just prior to, and 1 day after, noise exposure and 1 and 10 days after completing steroid administration. Each rat was anesthetized with Zoletil 50 (Virbac Laboratories, Carros, France) 0.1 cc/100 g and Rompun 2% (Bayer Korea, Ansan, Korea) 0.02 cc/100 g by intraperitoneal injection for auditory evaluation. ABR was recorded in a soundproof and electrically shielded room. Auditory evaluation was tested by measuring ABR with a Biosig 32 system (Tucker - Davis Technologies, Gainesville, FL, USA). Recording needle electrodes were inserted subcutaneously into the vertex (+) of the scalp and the postauricular area (−). Ground needle electrodes were inserted into the neck contralateral to the recording electrodes. The stimuli were tone bursts 5 ms in duration with rise/fall times of 1 ms. The sound frequencies delivered were 16 and 32 kHz to the left ear at a rate of 11.1/s. Responses were averaged over at least 512 repetitions of stimuli, which were decreased by 5 dB at subthreshold levels. The thresholds were defined using visual analysis by two investigators blinded to the noise + DEX group.
Histologic analysis
The extracted cochleae were placed in 4% (v/v) paraformaldehyde in phosphate-buffered saline (PBS) overnight and decalcified in Calci-Clear Rapid (National Diagnostics, Charlotte, NC, USA) for 3 days. After decalcification, the tissues were embedded in paraffin blocks, and sections 5 μm thick were cut along the cochlear axis and stained with hematoxylin and eosin (H&E). Hair cells and spiral ganglion in the cochlea were observed.
Immunohistochemistry
The sections were rehydrated. After washing with PBS containing 0.05% Triton X-100 (Wash buffer), blocking with 1% bovine serum albumin for 1 hour, and washing with Wash buffer, the sections were incubated with anti-C-terminal binding protein 2 antibody (CtbBP2, 1:300 dilution; BD Biosciences, Franklin Lakes, NJ, USA) and anti-Myosin VIIa antibody (MYO7A, 1:300 dilution; Cell Signaling Technology, Danvers, MA, USA) at 4°C overnight. After washing in Wash buffer, the sections were incubated with fluorescein isothiocyanate - conjugated secondary antibodies (1:300 dilution in PBS containing 0.2% Triton X - 100 [PBST]; Jackson ImmunoResearch, West Grove, PA, USA) and Cy™3 - conjugated secondary antibodies (1:600 dilution in PBST; Jackson ImmunoResearch) for 1 hour. After washing with Wash buffer three times for 5 minutes each time, the sections were mounted with VECTASHIELD mounting medium with DAPI (Vector Laboratories, Burlingame, CA, USA). The sections were examined using a Zeiss LSM510 confocal microscope (Carl Zeiss Microscopy, Jena, Germany).
H&E staining and immunohistochemical staining of the normal cochlea of SD rats (8 weeks old, 250–300 g) were performed for comparison with the noise and noise + steroid groups.
Data analysis
All data are presented as means ± standard error. The sample size was small in each group, so the values of hearing threshold and GPIAS were analyzed using the Mann-Whitney U test. In all analyses, P < 0.05 was taken to indicate statistical significance.
Ethics statement
All of the animals were managed in compliance with the Guidelines for the Care and Use of Laboratory Animals of the National Institutes of Health, and all of the experiments were performed in accordance with the Guidelines of the Internal Animal Care and Use Committee of Ajou University Hospital Biomedical Research Institute (protocol No. 2018-0005).
Go to :
RESULTS
ABR
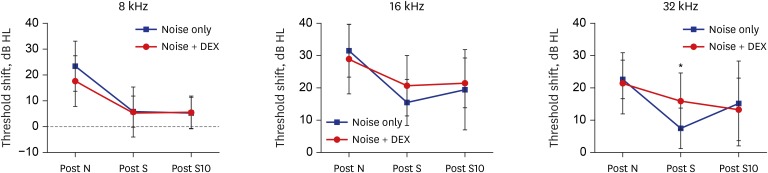
ABR was measured in all rats before noise exposure and 1 day after exposure to determine whether noise trauma could induce auditory threshold shift. Baseline threshold values in both groups were under 15 dB SPL at 8, 16, and 32 kHz. The threshold shift after noise exposure was not different between the two groups at 8, 16, and 32 kHz. Threshold shift was checked 1 and 10 days after intraperitoneal steroid injection, and showed no differences between the two groups in each frequency, except 32 kHz at 1 day after steroid injection (Fig. 3).
Tinnitus development and GPIAS
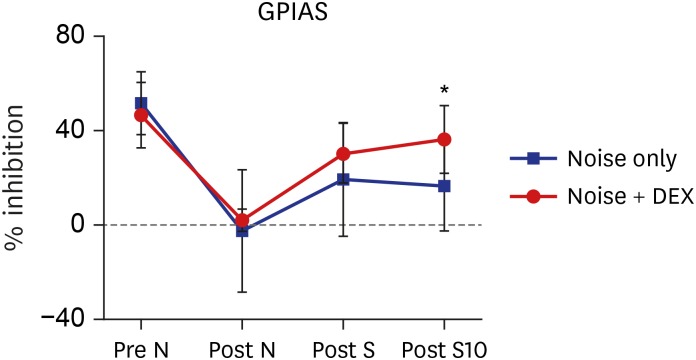
During baseline sessions, rats showed a mean GPIAS of 51.2% ± 13.4% in the noise group and 46.6% ± 14.1% in the noise + DEX group (P = 0.484). The mean GPIAS values did not show significant differences between the two groups at 1 day after noise exposure or 1 day after intraperitoneal steroid administration (P = 0.650 and P = 0.289). In contrast, the mean GPIAS value in the noise + DEX group (36.4% ± 14.1%) was significantly greater than that in the noise group (16.4% ± 18.8%) at 10 days after intraperitoneal steroid administration (P = 0.017) (Fig. 4).
Histology
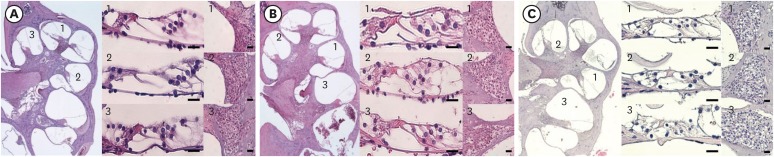
Histological analyses of the two groups were performed after functional evaluation. Fig. 5 shows the histological findings for the organ of Corti in the basal, middle, and apical turns. There were no pathological findings associated with noise trauma, such as loss of hair cells and supporting cells, in the two groups.
Immunohistochemistry
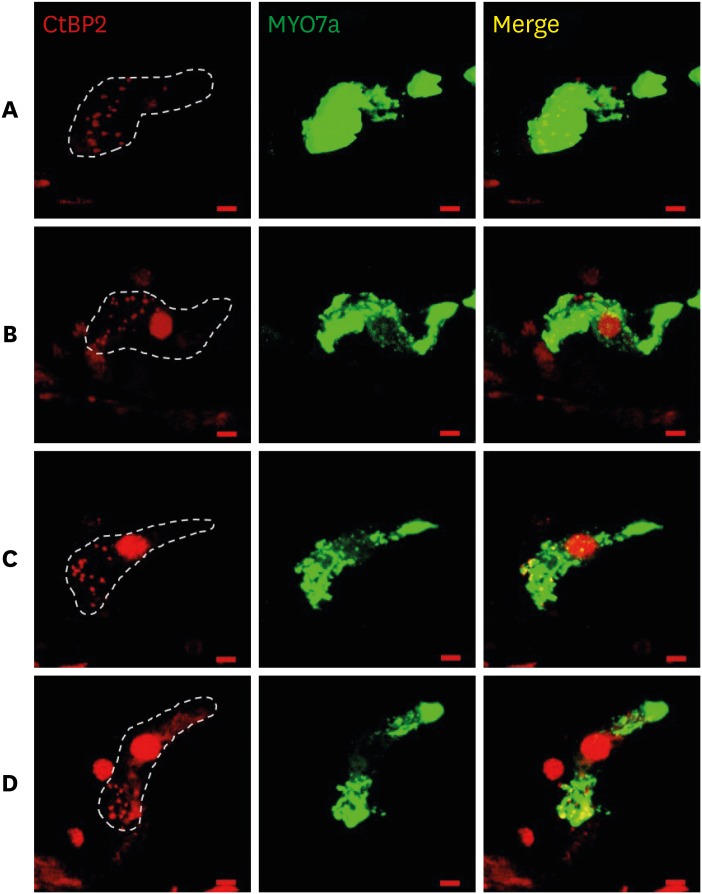
Immunostained cochlear sections, imaged by confocal microscopy, were assessed to quantify hair cells, cochlear neurons, and synaptic structures providing the communication conduits. Auditory nerve fibers (ANF) terminate in a single synapse on a single inner hair cell (IHC).16 Thus, the ribbon count provides an accurate metric of the number of ANFs contacting each IHC. The mean counts of ribbon synapse were similar in the noise group and noise + DEX group (Fig. 6).
Go to :
DISCUSSION
Various treatments have been used to control tinnitus, including tinnitus retraining therapy, medication, hearing aids, and transcranial magnetic stimulation. However, there have been few reports showing significant and long-lasting improvement of tinnitus.1718
Previous studies supported that damage to cochlear hair cells was essential for the pathogenesis of tinnitus.14 Therefore, in the initial stages of tinnitus, the period of reversibility of cochlear lesions, it was considered that acute tinnitus could be eliminated by appropriate therapy. As steroid receptors were identified in the inner ear,19 DEX has been used as an initial medication option for inner ear diseases and its effects on acute tinnitus have been investigated.202122 The efficacy of DEX used in clinic for the treatment of acute tinnitus was also supported by this animal study.
The present study was performed to evaluate objectively the effects of steroid administration on acute tinnitus in an animal model by measuring GPIAS. Since its introduction by Turner et al.,12 GPIAS has been widely used to investigate the mechanisms underlying tinnitus in animal models. The advantage of this method for tinnitus screening in rodents is that it does not require training and, instead, uses reflex modification procedures. The GPIAS system is based on the startle response, which is elicited by loud sounds. We checked the hearing level of the rats because hearing loss is known to affect the reliability of GPIAS by attenuating the behavioral response to loud sound perception. To measure the GPIAS value, it is essential to maintain the acceptable hearing level after noise exposure. Acoustic trauma is most commonly used to induce tinnitus in animal models. Galazyuk and Hébert10 summarized the tinnitus protocol of animal model. Sound levels vary from 94 dB SPL to 125 dB SPL. The most common parameters for tinnitus induction include narrow band noise (10, 12, 16, and 17 kHz) presented during 1–2 hours. Based on the study by Galazyuk and Hébert10 we attempted to create noise exposure conditions that caused tinnitus while preserving hearing as much as possible. The mean hearing thresholds of both groups were lower than the background noise (60 dB SPL) and startle stimulus (105 dB SPL) in the GPIAS measurement protocol. Tinnitus perception was shown to be significantly reduced in the noise + DEX group at 10 days after intraperitoneal steroid administration compared to the noise group by measuring the GPIAS value, which means tinnitus intensity was weakened by steroid. Acute insult such as noise trauma causes cochlear damage by reactive oxygen species accumulation23 and cochlear ischemia,24 which could initiate the tinnitus without hearing impairment. The mechanism of action of steroid on tinnitus is related with the decrease in inflammation due to immune system dysfunction in the inner ear, suppression of irritability or sensitivity due to damage of cochlear hair cells, and increased inner ear vascularity.25 These effects of steroid may contribute to tinnitus awareness control in rats with noise trauma.
We performed histological analysis of the noise and noise + DEX groups to compare the differences according to GPIAS value. No pathological changes were observed, such as the loss of hair cells and supporting cells, associated with noise trauma in the noise group and the noise + DEX group. The animal model in this study did not represent a model of tinnitus with normal auditory thresholds as hearing thresholds of 16 kHz increased to 40 dB HL. Sanchez et al.26 reported that only 7.4% of tinnitus patients showed normal pure tone thresholds. These findings suggested that acute tinnitus could be induced without structural damage to the cochlea and the animal tinnitus model with normal hearing category might be developed by adjusting the noise exposure condition.
A recent study showed that immediate and extensive loss of synapses between IHC and ANF can be caused by titrated noise exposure, which is designated as “synaptopathy”.27 We performed immunohistochemical staining to investigate the synapses of IHC in the present study. Although both the noise and noise + DEX groups showed similar patterns of staining for CtBP2, a presynaptic marker protein associated with synaptic ribbons, it should be considered that this finding could be rather preliminary using a small number of samples and ribbon counting in cochlear whole mount specimens and measurement of ABR wave I amplitude should be performed to confirm synaptopathy.
The clinical results regarding the effects of steroid administration on tinnitus control are controversial.2228 In this study, tinnitus control using steroid administration was validated in a model of acute tinnitus caused by acoustic trauma, supporting the positive effect of steroids.29 Steroids may eliminate the reversible inflammatory changes in the cochlea at the initial stages of tinnitus.
There are several factors to be considered. The effect of intratympanic DEX injection, commonly used in clinical practice, on acute tinnitus should be verified. Ribbon counting data in cochlear whole mount would be helpful. Additionally, comparison with the experiment using salicylate-induced tinnitus model might be significant.
In conclusion, we have developed a model of acute tinnitus with minimal structural change associated with noise exposure and objectively verified tinnitus by measuring the GPIAS value. Steroid therapy for tinnitus control was validated in this animal model. Additional trials for other tinnitus models associated with ototoxicity and sudden hearing loss could be required.
Go to :
 XML Download
XML Download