

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Respiratory syncytial virus (RSV) is one of the most common pathogens causing lower respiratory tract infections (LRTIs) in infants and young children. In Korea, the seasonal RSV epidemic usually begins in September, and the incidence peaks during December to January. Most cases occur in children below 6 years old.1 RSV infections present with a broad range of symptoms, from mild cough and rhinorrhea to severe wheezing and respiratory distress. In high risk children who are born premature, or in patients with chronic lung disease, congenital heart disease, or compromised immunity, RSV infections may lead to respiratory failure.2
RSV is a non-segmented negative strand RNA virus of the Paramyxoviridae family, Pneumovirus genus.3 It contains 10 genes that encode 11 proteins and the genome is approximately 15.2 kb in size. The RSV protein is composed of a nucleocapsid protein (N) that protects the RNA genome, phosphoprotein (P), matrix protein (M), which forms the viral membrane, glycoprotein (G), which binds to cells, fusion protein (F), which fuses to the virus and infects the cell after attachment to cell, RNA polymerase large protein (L) for RNA replication, membrane proteins 2-1, 2-2 (M2-1, M2-2) which mediate transcriptional regulation, small hydrophobic protein (SH), and non- structural protein 1, 2 (NS1, NS2).4 Of these, F and G proteins are the major surface glycoproteins and the antigenic difference between these two proteins divides RSV into two genetically different subgroups RSV-A and RSV-B.5
While F protein does not show much variation, G protein contains a hypervariable region, thus, the G gene that encodes the G protein is highly variable among RSV strains. The RSV gene consists of about 15,000 nucleotides in total, of which 1,000 constitute the G gene and can be found at positions 4,500–5,500, approximately.
The emergence of novel genotypes due to G gene variations can sometimes cause recurrent RSV infection6 and serious pandemics.7 Recently, the RSV-A ON-1 genotype has been increased worldwide. Therefore, this study aimed to identify the prevalent genotypes in Korea through analyses of the G gene in RSV-A and RSV-B strains that were prevalent in during the 2017 and 2018/2019 seasonal epidemics, and observe changes in the epidemiology of circulating RSV genotypes and genetic variability of G gene in Korea.
METHODS
Subjects
Patients below 14 years old that were admitted to the pediatrics department at the Catholic University of Korea Daejeon St. Mary's Hospital during the 2017 and 2018/2019 RSV season for acute LRTI and tested positive for RSV in the multiplex polymerase chain reaction (PCR) obtained from the nasopharyngeal aspirates or nasal swab were included. The samples from the 2017 RSV season were collected during November to December 2017, and October 2018 to April 2019 in the 2018/2019 RSV season.
Viral nucleic acid extraction
All nasal specimens containing RSV RNA were stored at −70°C until the end of the study period. The viral nucleic acids were then extracted from the specimen using Ribospin™ vRD plus (GeneAll Biotechnology, Seoul, Korea) according to the instructions of the manufacturer.
Reverse transcription (RT)-PCR analysis
The extracted viral RNA was then used as a template for RT-PCR using Reverse Transcription Master Premix (Elpis Biotech, Daejeon, Korea) to synthesize cDNA. RT-PCR was made in a 20-uL reaction volume with 16-uLcontaining 100 or 500 ng of viral RNA and 10 pmol/uL of primers G151-173F (5′-CTGGCAATGATAATCT CAACTTC -3′) and F3-22R (5′-CAACTCCATTGTTAT TTGC-3′) each, 4 uL of virus reverse transcription master premix (Elpis biotech, Daejeon, Korea) containing 2 uL of 10× PCR Buffer, 2.5 mM of dNTP mixture 0.2 uL (10 U) of Taq DNA polymerase. The amplification was done for 40 cycles, comprising of 30 seconds at 94°C, 30 seconds at 55°C, and 45 seconds at 72°C, followed by 5 minutes of extension at 72°C. Extreme precaution measures were taken to prevent contamination during cDNA synthesis and PCR.
Gel electrophoresis
Ten microliters of the PCR product were analyzed by gel electrophoresis on 1% or 1.5% agar gel using a submarine-type electrophoresis device, Mupid-exU (Takara Bio Inc., Shiga, Japan). The band located at around 850 bp was checked under UV light and photographed using Image analyzer (Geldoc XR Image System, BioRad, Thermo Fisher Scientific, Waltham, MA, USA). Afterwards, the distinct band formed at about 800–900 bp was excised and a gel extraction procedure was carried out using a commercialized gel extraction SV kit (MGmed, Seoul, Korea).
DNA sequencing
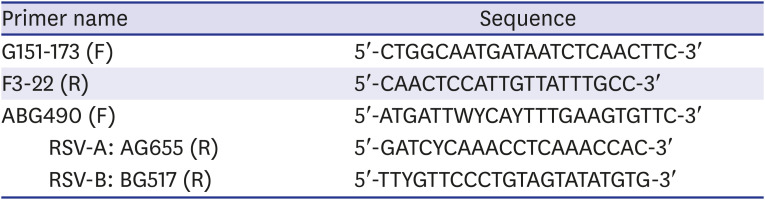
The DNA sequence of the gel-extracted cDNA was analyzed using BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Thermo Fisher Scientific, Foster City, CA, USA) and ABI PRISM 3730XL Analyzer (Applied Biosystem, Thermo Fisher Scientific). For DNA sequencing, four primers8 for each specimen were used (Table 1).
Phylogenetic analysis
When analyzing the G gene DNA sequence, multiple alignment was performed using Clustal Omega and the homology of the analyzed DNA sequence to the RSV G gene. DNA sequence registered to the GenBank database was checked using the National Center for Biotechnology Information Basis Local Alignment Search Tool (BLAST). For phylogenetic analysis, we first aligned the DNA sequences of previously reported reference strains from Genbank (Supplementary Tables 1 and 2) using a BioEdit version 7.0.5 and constructed trees using the neighbor-joining method with maximum composition likelihood algorithm in MEGA X software.5 The tree branch support values were measured by bootstrapping (1,000 iterations).
Glycosylation analysis
The glycosylation analysis was performed by predicting N-glycosylation sites using NetNGlyc 1.0 with a threshold value of 0.5.
Control RSV-A & B testing
RSV-A and RSV-B strains cultured in cells were purchased from the Korea Bank for Pathogenic Viruses and RT-PCR and gel electrophoresis assays were performed using the above-mentioned protocol. We confirmed band formation at around 800–900 bp and proceeded with testing of the corresponding specimens.
RT-PCR of the G gene was performed using the forward primer G151-173 and reverse primer F3-22 (Table 1).
Ethics statement
Written informed consent was waived because the clinical samples that the viruses were isolated from were taken for diagnostic purposes. The remaining samples were used for this study. The genetic sequencing of the viral RNA did not require written informed consent. This study was approved by the Institutional Review Board (IRB) of the Catholic University of Korea Daejeon St. Mary's Hospital (IRB No. DC18SESI0121, DC19SESI0038).
RESULTS
RSV seasonal outbreaks patterns
A total of 155 samples were positive for RSV: 49 cases (RSV-A, n = 5; RSV-B, n = 44) were positive for RSV in the multiplex PCR of the samples obtained from the nasopharyngeal specimens from the 2017 RSV seasonal epidemic, and 106 cases (RSV-A, n = 97; RSV-B, n = 9) from the 2018/2019 seasonal epidemic. During the two seasonal epidemics, RSV-B was dominant in the 2017 epidemic, while RSV-A was dominant in the 2018/2019 epidemic (Table 2). Of these, a total of 26 were included in the analysis of the hypervariable region of the G gene. Of these, 46.2% were male, and the mean age was 22 months old (standard deviation ± 28.9 months). The clinical diagnosis was acute bronchiolitis in 50.0% of the cases, and pneumonia in 42.3% (Table 3).
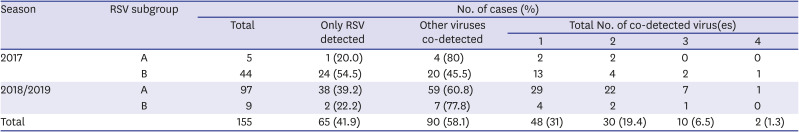
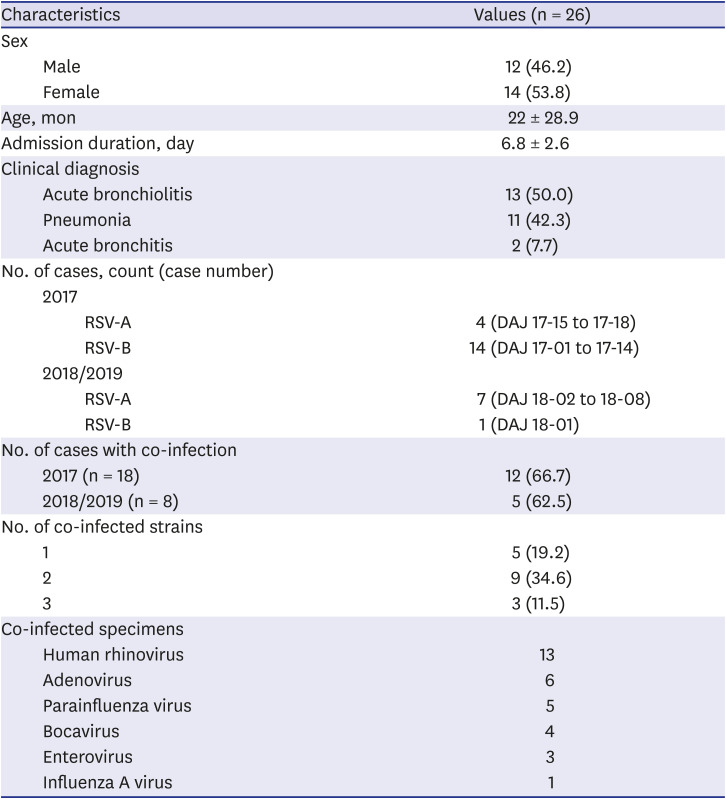
Table 2
Detection of RSV and other viruses in multiplex PCR
![]()
Table 3
Characteristics of subjects
![]()
In both seasonal epidemics, co-detection with viruses other than RSV were confirmed in more than 50% of the cases. In one third of these patients, one other virus was co-detected (Table 2). Human rhinovirus was the most common virus that was co-detected via real-time PCR test (Table 4).
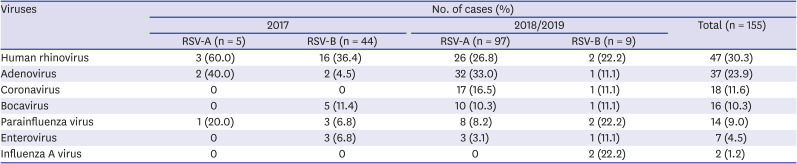
Table 4
Viruses detected in addition to RSV among specimens
![]()
A total of 18 cases (RSV-A, n = 4 [80%]; RSV-B, n = 14 [31.8%]) from the 2017 and 8 cases (RSV-A, n = 7 [7.2%]; RSV-B, n = 1 [11.1%]) from the 2018/2019 RSV seasonal epidemic were sequenced. Of the total 26 patients, 17 (65.4%) were co-detected with other virus; the percentage similar to that of the entire study population (Table 4). The mean age of the patients was 22 months, and the male-to-female ratio was 1:1. The patients were admitted for 4 to 10 days and the most common diagnosis was acute bronchiolitis (Table 3).
RSV genotypes circulating during RSV epidemics
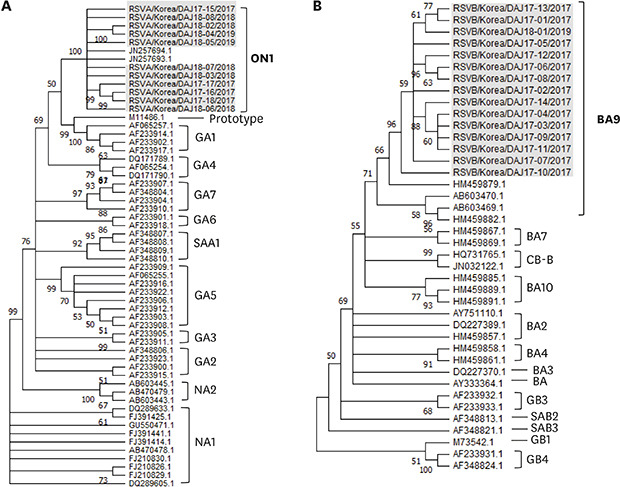
The sequences of the second hypervariable region of G gene was confirmed in 11 RSV-A and 15 RSV-B cases: 4 RSV-A and 14 RSV-B sequences from the specimens collected in the 2017 RSV seasonal epidemic, and 7 RSV-A and 1 RSV-B sequences from the specimens collected in the 2018/2019 epidemic. All sequences were referred to the G gene sequences already reported in the Genbank 4910 for comparison (Supplementary Tables 1 and 2). All RSV-A strains were classified into the ON1 genotype, while all RSV-B specimens were classified as the BA9 genotype (Fig. 1) during the two seasonal epidemics.
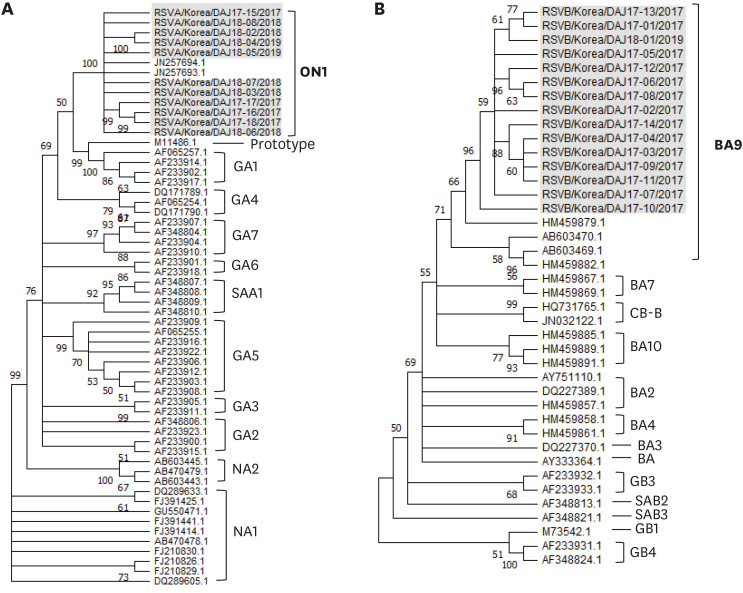
Fig. 1
Phylogenetic trees based on the second hypervariable region of the G protein. Trees using the neighbor-joining method with maximum composition likelihood in Mega X by bootstrapping (1000 iterations). Only bootstrap values ≥ 50% are shown for each tree. Our stains were expressed as name of strains/country/isolated case number/collected year. The control strains used for comparison were expressed as accession numbers registered in the Genbank. (A) RSV-A (All cases are classified as ON1 genotype, n = 11), (B) RSV-B (All cases are classified as BA9 genotype, n = 15).
RSV = respiratory syncytial virus.
![]()
The strains isolated in this study were compared with the enrolled G gene sequences of RSV via the MegaBLAST program to optimize for highly similar sequences. The sequences of the 25 strains were unprecedented, whereas only one strain (RSV-B, 2017) was identical to previously enrolled strains from Korea, 2018. Using RSV G gene sequences with the highest similarities at the nucleotide level for each of our strains, a phylogenetic analysis was carried out (Fig. 2). A subset (n = 2, DAJ18-02 and DAJ18-04) of RSV-A formed cluster with strains from Korea, 2015, 2017 and Tailand, 2016. However, RSV-A strains did not form a cluster as a whole, but rather partially formed a cluster and revealed high similarities to the strains reported over various countries and periods. One strain (DAJ18-06) formed a cluster with a strain from Korea, 2011. Three (DAJ17-16, DAJ17-17. DAJ17-18) of four RSV-A strains from 2017 formed a cluster together (Fig. 2). On the other hand, majority of RSV-B strains showed more similar genotypes with neighboring countries in the East Asian regions, such as China, Japan, Taiwan and other local regions of Korea. It also showed high similarities to strains found at similar periods (2016–2019). However, strains that caused epidemics in Switzerland and Russia during the 2018/2019 season showed high similarities with RSV-B specimens from our study.

Fig. 2
Phylogenetic trees based on the second hypervariable region of the G protein with genes with high similarity (97%–100% identity) via the MegaBLAST program. Trees using the neighbor-joining method with maximum composition likelihood in Mega X. Bootstrap values were 1,000 iterations. Only bootstrap values ≥ 50% are shown for each tree. Our stains were expressed as name of strains/country/isolated case number/collected year. The control strains used for comparison were expressed as accession numbers registered in the Genbank and strains/country/isolates/number/collected year. (A) RSV-A, (B) RSV-B.
RSV = respiratory syncytial virus.
![]()
Variability of amino acid sequences in the second hypervariable region of G gene of ON1 and BA9
The amino acid sequences of the strains were aligned and compared to reference ON1 (Fig. 2A) and BA9 (Fig. 2B) strains showing high similarities through MegaBLAST. The strains of RSV-A and RSV-B during the two outbreak seasons were examined for the region spanning residues 209–310 of G protein of ON1 (JN257693) reference strain and the region spanning residues 230–311 of G protein of BA (AY333364) reference strain, respectively (Fig. 3). Both ON1 and BA9 amino acid sequences of the original and duplicated regions showed that within one season, different mutations were observed. Also, when comparing the two RSV epidemic seasons, strains with novel mutations were observed.
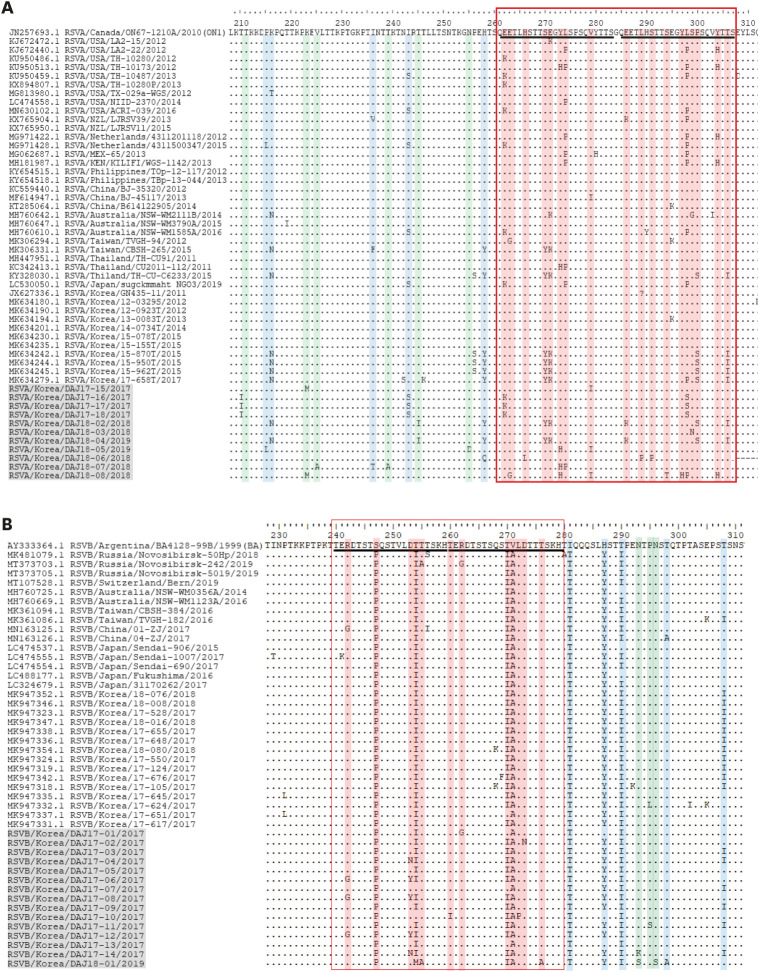
Fig. 3
Deduced amino acid alignments of the second variable region of the G genes. The amino acid positions corresponded to amino acid positions 210 or 230 to 310 of the reference sequence. The original and duplicated 23-amino-acid region in RSV-A group and 20-amino-acid region in RSV-B group are indicated by rectangles. Our stains were expressed as name of strains/country/isolated case number/collected year. Control strains used to compare were expressed as accession numbers registered in the Genbank and strains/country/isolates/number/collected year. (A) RSV-A (The representative RSV-A ON1 genotype strain was ON67-1210A, GenBank accession number JN257693); (B) RSV-B (The representative RSV-B BA genotype strain was BA4128/99B, GenBank accession number AY333364).
RSV = respiratory syncytial virus, blue color = the sequence mutated found in other strains which were previously reported, green color = the sequence mutated relatively new compared to strains with high similarities, red color = the sequences mutated in the duplicated region.
![]()
At the amino acid level of the second hypervariable region of RSV-A G protein, three stains (DAJ17-16, 17-17, 17-18) had identical nucleotide substitutions (T210I, I243S, E262K, L298S). Two strains (DAJ18-02, 18-04) had the same changes of K216N, T245I, H258Y, S270Y, E271K, E286K, P300S, T206I in the amino acid sequences. The remaining RSV-A strains each had independent mutations in the duplicated region compared with the amino acid sequences of the Ontario ON1 strain (JN257693, RSVA/Canada/ON67-1210A/2010) (Fig. 3). N-glycosylation sites were investigated and only one strain (DAJ 18-3) had one N-glycosylation site in the range of the duplicated region (261-298 position of aa in G-protein) due to the substitution of S299N.
All RSV-B strains showed V271A substitution which is characteristics in BA9 subtype. Three strains (DAJ17-06, 17-08,17-12) had identical mutations such as R242G, D253Y, T270I in the duplicated amino acid region (Fig. 3). Two strains (DAJ17-04, 17-14) and one strain (DAJ17-02) showed D253N and D273N, predicting a higher number of N-glycosylation sites. Especially in the 2018/2019 season, BA-9 strain (DAJ18-01) showed different unique amino acid sequences with T254M, T276A, N293S, N296S, T298A mutation which did not appear in the strains from 2017. Furthermore N-glycosylation site at position 296, which had known to be conserved in all BA viruses, were substituted with ‘S’ in this case. Our RSV-B strains tended to have fewer predicted N-glycosylation sites than RSV-A at positions 180–320 of the G protein. In addition, DAJ18-01 strain had only one predicted N-glycosylated site, which was the smallest number of N-glycosylation sites in our cases (RSV-A and RSV-B).
DISCUSSION
The present study investigated the trends and genotypes of the RSVs that were prevalent in the 2017 and the 2018/2019 seasonal epidemic in the Daejeon area. The study showed that the RSV-B BA 9 genotype was dominant in 2017 epidemic, and the RSV-A ON1 genotype was dominant in the 2018/2019 epidemic in Daejeon. Except for one RSV-B strain (DAJ17-10), which showed 100% homology with another Korean strain previously reported,11 all the G gene nucleotides of the RSVs in this study had novel sequences.
The G protein encoded by the G gene is a type II glycoprotein composed of about 300 amino acids that contains a hypervariable region at one-third of the C-terminal region of the ectodomain. Such characteristic makes it possible for different G protein subgroup genotypes to be expressed based on differences in antigen and genomic sequence, some of which have been reported 18 RSV-A genotypes (GA1-7,12 SAA1-2,12 NA1-4,13 CB-A,9 and ON1-4,1415) and 35 RSV-B genotypes (GB1-13,1216 BA1-14,1017 CB-B,18 SAB1-4,19 THB,20 URU1-221) and the number continues to increase.
RSV-A ON1 genotype was first reported in Ontario, Canada, in 2011. The ON1 genotype was found to have a 72-nucleotide duplication in the C-terminal region of the G gene. ON1 genotype tends to replace NA1, as reported worldwide. In Korea, Baek et al.9 reported that the NA1 genotype was dominant among RSV-A cases in 2008–2009 and 2009–2010, while Hwang et al.22 reported that the RSV-A ON1 genotype was detected in 20 cases in 2011–2012 (74.15%), 23 in 2012–2013 (95.8%), 1 in 2013–2014 (100.0%) and 9 in 2014–2015 (100%). Since the ON1 genotype first emerged in Korea in 2011522 only the ON1 genotype has been reported in RSV-A since 2014.2223 In this study, all 11 RSV-A from the 2017 and 2018–2019 seasons were confirmed to be of the ON1 genotype. This suggests that the RSV-A NA1 genotype prevalent in Korea has been replaced by ON1.
In 1998, in Madrid, a 60-nucleotide duplication of the BA genotype was first discovered in the second hypervariable region of G protein, which was first named the BA genotype in 1999, in Buenos Aires.10 The BA genotype is prevalent worldwide and has been reported in Japan,24 Kenya,25 Belgium,26 Canada, Brazil, the United Kingdom and the United States27 and has been reported to be the most dominant genotype among RSV-B since 2006,28 which may be related to the fact that a 60-nucleotide duplication causes an antigenic change in the G protein, allowing it to evade host immune response.27 Thus far, the BA genotype is divided into 12 subtypes, from BA1 to BA12, and the BA9 genotype was first found in 2006–2007 in Japan.29 It later became prevalent in 2009–2010 in Japan and was also reported to be the major genotype in the Philippines from 2009 to 2010.30 In China, BA9 started to become prevalent from 2006 to 2011.31 When BA9 was first classified and named in Japan, the BA4 branch was re-classified into BA7,8,9,10. Recent studies have reported that BA9 and BA10 prevalence decreased from 2010 to 2014 in Japan, whereas in China BA10 prevalence tended to decrease and that of BA9 increased over the same period. Yun et al. underwent an extensive investigation of the circulating RSV genotypes during 28 seasons from 1990 to 2018 in Korea. they showed that RSV-A ON1 genotype and RSV-B BA9 genotype have been the dominant circulating genotypes during the past decade. In that study, RSV-B was dominant during 1998/1999, 2005/2006, 2010/2011, 2013/2014, 2015/2016, and 2017/2018 epidemics compared to RSV-A and BA genotype was appeared from 2005/2006. BA 9 was replaced predominantly from 2008/2009 and replace completely from 2015/2016 epidemics with shortening the epidemic cycle of RSV-B.11 Although the region of the study was different from this present study, this study showed that BA9 RSV-B was prevalent during 2017/2018 epidemics while RSV-A infection was prevalent from fall of 2018 to the following spring in Korea.
None of the strains of RSV-A had identical sequences of the duplicated region of the G protein as the Ontario strain (JN257693) (Fig. 3). However, three specific substitutions (E232G, T253K, and P314L) only observed from Ontario14 were seen in all RSV-As in the present study. Amino acid sequences of nine among eleven strains of RSV-A were newly reported and only two strains (DAJ17-15, 18-7) showed identical sequences in the duplicated regions of strains of 2016/2017 and 2017/2018 from Yun et al.'s11 study. Unique sequences of amino acids such as such as E263G (DAJ18-08), H266L (DAJ18-07), L289P (DAJ18-06), S291P (DAJ18-06), S294T (DAJ18-08), Y297H (DAJ18-08), L298S (DAJ17-16,17-17,17-18), S299N (DAJ18-3), Y304H (DAJ18-8) which have not been reported in Korea were found in strains from the 2018/2019 season. This is the first report of latest strains in Korea. From 2017, three (DAJ17-16,17-17,17-18) out of four strains formed a cluster, and a novel sequence that has not been previously reported in Korea were identified. It suggests that genetic evolution occurs variously by regional differences even during the same period and within the same country.
In the BA9 genotype, it has been reported that the V amino acid in position 271 was substituted to A and similarly in the present study, all RSV-B showed the V271A substitution and N-glycosylation was found in the amino acid positions 296 and 310, which is in line with previous findings (Fig. 3).10 The location of the N-glycosylation site at amino acid position 310 was conserved across all RSV-B specimens. Four strains (DAJ17-4, 17-10, 17-14, 18-01) of fifteen strains of RSV-B showed novel sequences in duplicated regions in Korea. In addition, T254M (DAJ18-01), T255A (DAJ18-01), T260I (DAJ17-10) was first reported in Korea. The one strain from 2018/2019 had a new sequences and novel substitution in duplicated region compared to them of 2017, which can be attributed to genetic evolution of strains every year.
RSV-A showed high similarity with strains previously reported during the past decade, regardless of regional proximity. This may be because RSV-A is more consistently the cause of epidemics worldwide, as well as nationwide epidemics than RSV-B. On the other hand, RSV-B strains showed high similarity to strains in close proximity and recent epidemic seasons. Recent BA strains which showed high similarity with our strains tended to have S247P, T254I, T270I, V271A, I281T, H287Y, T290I in second hypervariable region of G protein compared Buenos Aires (AY333364) (Fig. 3). Especially, strains from Korea, including our strains, had a tendency of having T308I mutations recently (Fig. 3). This suggests that even though RSV-B has fewer mutations than RSV-A, genetic mutations develop over time regionally and regularly. Thus, it should be taken into account that though the pathogenicity of BA9 may be strong enough to drive the outbreak of RSV-B rather than RSV-A, the host's defense system against the BA9 mutation and rapid genetic mutation of RSV-A may cause low frequency of domestic outbreaks compared to RSV-A.
There were several limitations in this study. First, this study was conducted during a short period of 2017 and 2018/2019 RSV season at a single center in Daejeon. Although the number of strains included were small, it represents the genotypes of a region in Korea. Second, because the RNAs of the viruses were collected from specimens after testing in patients without RSV cell culture, the yield was low. However, the accurate gene analysis of RSV, including the hypervariable region was possible using this method.
The present study is the only study reported thus far, which shows that RSV-B of the BA9 genotype was prevalent in 2017. It also showed that RSV-A that was prevalent in the 2018/2019 season was of the ON1 genotype. By identifying subtypes and genotypes of RSV that are prevalent in local region, it can provide information to understand the overall RSV epidemic pattern in Korea. Furthermore, it will be useful to confirm the correlation between region and clinical progress in the aspect of genetic evolution of RSV in the future which is significant for clinical and public health implications.
 XML Download
XML Download