

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Estrogen is known to have two opposite effects on the growth of longitudinal bones. It increases growth by increasing growth hormone (GH) and insulin like growth factor-I (IGF-I) secretions during the pubertal growth spurt, but it stops growth by promoting the closure of the growth plates with differentiating chondrocytes. It also promotes the accretion of bone mineral density (BMD) through the differentiation of osteoblasts and osteoclasts.1
In the classical pathway, the effects of estrogen occur after binding to estrogen receptors (ER, ESR). Two subtypes of ER have been identified, estrogen receptor α (ERα, ESR1) and β (ERβ, ESR 2).2
If each ER subtype has different effects on regulating pubertal onset, growth spurts, growth plate closure, acquisition of bone mineral content, and reproductive functions, we may control these actions by manipulating the estrogen binding intensity to each ER subtype. We might be able to increase the final height in adults without markedly reducing BMD or impairing reproductive functions, even in humans.3456
Nowadays, the incidence of precocious puberty in children is increasing in developed countries. It causes short stature due to earlier pubertal growth spurt and closure of epiphyseal plates in long bones.7 If we can elucidate the action of each ER subtype on pubertal growth spurt and epiphyseal plate fusion, we may find out the method to increase human height more efficiently.
The aim of our study was to understand the actions of each ER subtype on the pubertal growth spurt, growth plate closure, and acquisition of bone mineral content.
Go to : 
METHODS
Animals
Thirty female Sprague Dawley rats were housed in an approved animal facility with a 12-hour light cycle and given ad libitum access to food and water. They were ovariectomized at 4 weeks of age under intramuscular and intraperitoneal anesthesia with 30 mg/kg of tiletamine hydrochloride (HCl) and zolazepam HCl (Zoletil® 10%; Virbac, Carros Cedex, France) and 5 mg/kg of xylazine HCl (Rompun® 2%; Bayer, Leverkusen, Germany).
Drug injections
The rats were randomly divided into 3 groups (n = 10/group). Ten rats were injected with sesame oil (control group); another 10 were injected with 10 mg/kg of propylpyrazole triol (PPT®; Cayman Chemical, Ann Arbor, MI, USA) as the ERα agonist treated group (PPT group); the other 10 were injected with 10 mg/kg of 2,3-bis (4-hydroxyphenyl) propionitrile (DPN®; Cayman Chemical) as the ERβ agonist treated group (DPN group). The rats were subcutaneously injected with the same volume starting at the age of 6 weeks, 5 days per week for 5 weeks.
Body length and weight measurements
The body length (crown-rump length) and weight of each rat were measured weekly from the age of 1 through 10 weeks.
BMD analysis
Analyses of total body and lumbar vertebral BMDs of the rats were performed at the age of 6 and 10 weeks through dual energy X-ray absorptiometry (DXA) using the Lunar PIXImus mouse densitometer (Wipro GE Healthcare, Madison, WI, USA), the Norland Medical systems pDEXA Sabre (Norland Medical Systems, Fort Atkinson, WI, USA), and the Sabre Research Software (version 3.6; Norland Medical System).
Measurement of serum hormone levels
Blood samples were obtained via the tail vein of 6- and 10-week old rats, and serum levels of GH and estradiol (E2) were determined with the ELISA kits for GH (Millipore, Darmstadt, Germany) and E2 (Calbiotech, Spring Valley, CA, USA).
Quantitative real-time polymerase chain reaction (RT-PCR) analysis
After each rat was euthanized, Gh1 mRNA was extracted from the pituitary gland with RNeasy mini kit (Qiagen, Duesseldorf, Germany) according to the manufacturers' instructions. The quantitative RT-PCR analysis was performed using the ABI Power SYBR green PCR master mix (Thermo Fisher Scientific, Waltham, MA, USA) and the Step One Plus RT-PCR system (ABI). The sequences of the primer sets used for Gh1 and 18S were as follows; sense 5′-GCTGCAGACTCTCAGACTCCCTGG-3′, antisense 5′-CTGAGAAGCAGAACGCAGCCTG-3′, sense 5′-TGGTTGATCCTGCCAGTAG-3′, and antisense 5′-CGACCAAAGGAACCATAACT-3′.
Quantitative histology of growth plates
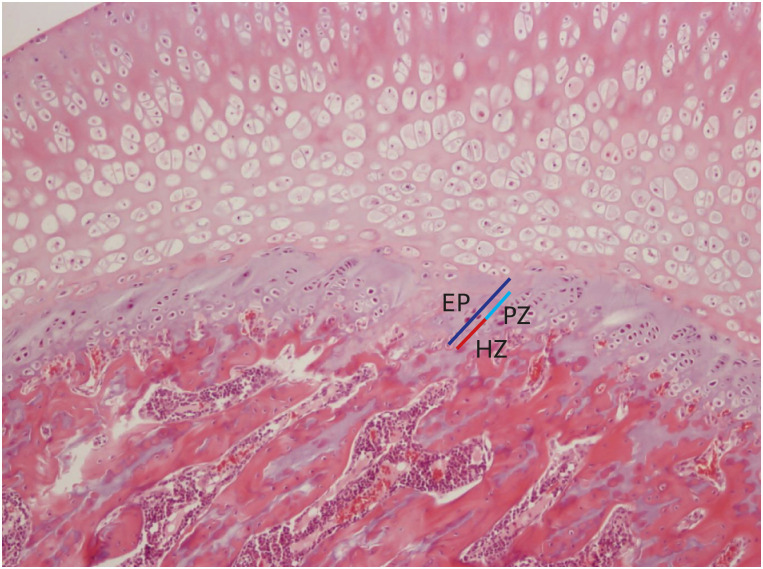
The proximal tibias of the rats were dissected, decalcified, and stained with hematoxylin-eosin. The thicknesses of their epiphyseal plates (EP) including the proliferative (PZ) and hypertrophic zones (HZ) were determined on 20-evenly divided sites in the central three-fourths of the growth plate sections using a Nikon Eclipse E800 light microscope (Nikon, Tokyo, Japan) with ImageJ software (version 1.5, NIH, USA) (Fig. 1).8
Statistics
The differences in auxological data, BMD, serum GH and E2 levels before and after injections were analyzed with the one-way analysis of variance with multiple comparisons. The Kruskall-Wallis test was used to compare the pituitary Gh1mRNA levels and histologic data among the groups with the SPSS ver. 20.0. All the data were expressed as mean ± standard deviation. P < 0.05 was considered as statistically significant.
Ethics statement
The procedures used and the care of animals were approved by the Institutional Animal Care and Use Committee in the Kyung Hee University Hospital at Gangdong (approval No. KHNMC AP 2013-011).
Go to :
RESULTS
Change of auxological data
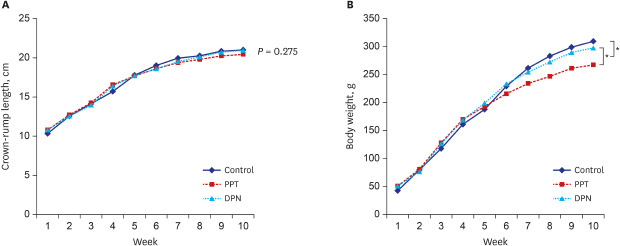
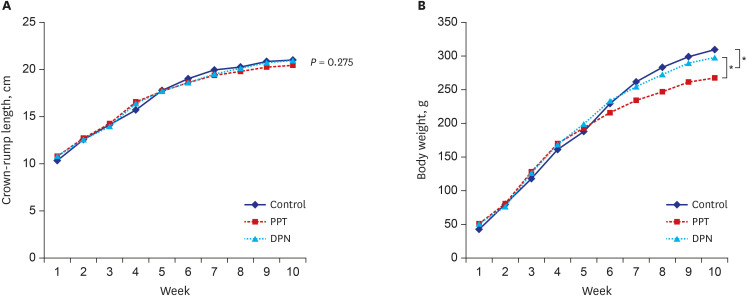
The body lengths of 1-week-old rats in the control, PPT, and DPN groups were 10.33 ± 0.75, 10.79 ± 0.51, and 10.8 ± 0.5 cm (P = 0.275). At 4 weeks of age and before the injections, the body lengths were 15.7 ± 0.68, 16.56 ± 0.53, and 16.39 ± 0.49 cm for each group (P = 0.969); and at 10 weeks of age and after injections 21.0 ± 0.52, 20.44 ± 0.29, and 20.94 ± 0.4 cm for each group (P = 0.083). There was no significant difference in body length among 3 groups before and after injections (P = not significant).
The body weights of 1-week-old rats in the control, PPT, and DPN groups were 42.52 ± 3.11, 50.85 ± 4.78, and 50.92 ± 4.48 g (P = 0.172). At 4 weeks of age and before injections, the body weights were 161.05 ± 10.71, 169.94 ± 15.3, and 168.54 ± 8.19 g for each group (P = 0.637); and at 10 weeks of age and after injections, 309.46 ± 22.65, 267.49 ± 16.82, and 297.66 ± 16.25 g for each group (P = 0.012). The mean body weight after 5 weeks of injections in the PPT group was significantly lower than that in the other 2 groups (P < 0.05) (Fig. 2).
Bone mineral density
The total body BMD in the control, PPT, and DPN group at 6 weeks of age was 0.101 ± 0.004, 0.108 ± 0.008, and 0.105 ± 0.008 g/cm2 (P = 0.083), and at 10 weeks of age was 0.121 ± 0.006, 0.146 ± 0.009, and 0.124 ± 0.009 g/cm2 (P = 0.003). The lumbar vertebral BMD in the control, PPT, and DPN group at 6 weeks of age was 0.093 ± 0.001, 0.099 ± 0.001, and 0.098 ± 0.002 g/cm2 (P = 0.075), and at 10 weeks of age was 0.135 ± 0.006, 0.155 ± 0.009, and 0.138 ± 0.009 g/cm2, respectively (P = 0.038). Therefore, the total body and lumbar vertebral BMD were significantly increased in PPT group than those in the other 2 groups (P < 0.05) after 5 week-injections (Table 1).
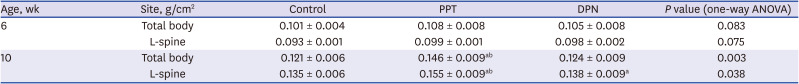
Table 1
The changes of bone mineral density before and after injections in three groups
Data are expressed mean ± standard deviation.
L-spine = lumbar spine, PPT = propylpyrazole triol, DPN = 2,3-bis (4-hydroxyphenyl) propionitrile, ANOVA = analysis of variance.
aP < 0.05 vs. control; bP < 0.05 vs. DPN.
![]()
Serum hormone levels
There was no significant difference in serum levels of GH and E2 in 6-week-old rats among 3 groups. The serum GH level in 10-week-old rats was 3.36 ± 0.18, 7.29 ± 0.58, and 3.84 ± 0.37 pg/mL in the control, PPT, and DPN groups, respectively. Therefore, the serum GH level in the PPT group was significantly increased compared with those in the other 2 groups (P < 0.05). The serum E2 level in 10-week-old rats was 6.11 ± 0.92, 5.45 ± 1.38, and 5.94 ± 1.23 µg/mL in the control, PPT, and DPN groups, respectively. There was no significant difference in serum E2 levels among the groups (P = 0.602) (Table 2).
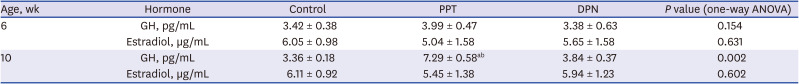
Table 2
The changes of serum hormone levels before and after injections in three groups
Data are expressed as mean ± standard deviation.
GH = growth hormone, PPT = propylpyrazole triol, DPN = 2,3-bis (4-hydroxyphenyl) propionitrile, ANOVA = analysis of variance.
aP < 0.05 vs. control; bP < 0.05 vs. DPN.
![]()
Gh1 expression levels in the pituitary gland
The relative expression level of the Gh1 gene was 0.94 ± 0.14, 1.44 ± 0.66, 1.23 ± 0.2 in the control, PPT, and DPN groups, respectively. The Gh1 expression was significantly increased in the PPT and DPN group (P < 0.05) (Fig. 3).
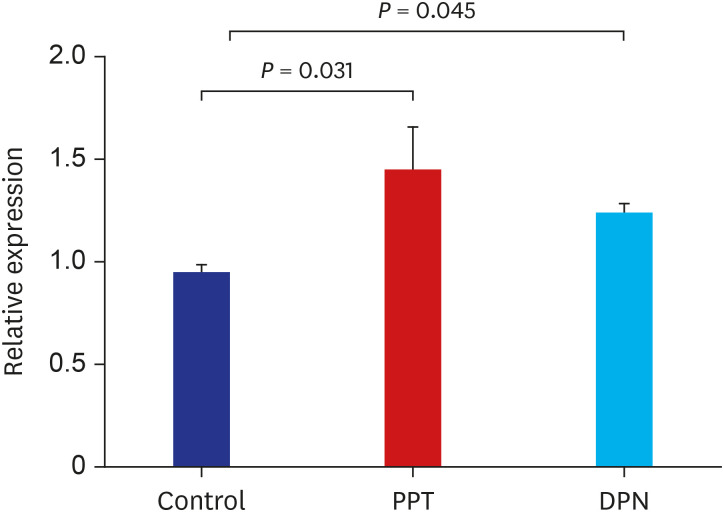 | Fig. 3Relative expression of Gh1 gene in the control, PPT, and DPN groups of rat after 5 weeks of treatment with sesame oil, PPT, or DPN. The expression was determined using quantitative RT-PCR.PPT = propylpyrazole triol, DPN = 2,3-bis (4-hydroxyphenyl) propionitrile, RT-PCR = real-time polymerase chain reaction.
|
Quantitative histology
The thicknesses of the proliferative zones were 45.77 ± 1.7, 45.12 ± 2.98, and 41.78 ± 1.2 μm in the control, PPT, and DPN group after injection, respectively, and there was no significant difference among the groups (P = 0.332); those of the hypertrophic zones were 35.48 ± 2.09, 34.4 ± 1.62, and 30.58 ± 1.03 μm, respectively, and there was no significant difference among the groups (P = 0.226). The thicknesses of total epiphyses were 91.25 ± 2.41, 89.51 ± 2.66, and 82.36 ± 2.85 μm, respectively, and there was no significant difference among the groups (P = 0.251) (Fig. 4).
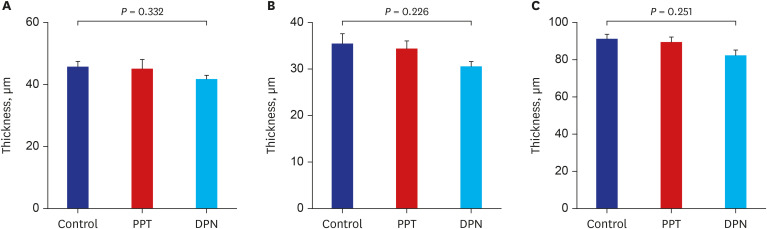 | Fig. 4Comparison of the epiphyseal plate thicknesses including proliferative and hypertrophic zones among 3 groups of rat after 5 weeks of treatment with sesame oil, PPT, or DPN. (A) thickness of the proliferative zone (B) thickness of the hypertrophic zone (C) thickness of the total epiphyseal plate.PPT = propylpyrazole triol, DPN = 2,3-bis (4-hydroxyphenyl) propionitrile.
|
Go to :
DISCUSSION
Estrogen affects the growth, differentiation, and development of a broad range of target tissues, such as those of the reproductive, skeletal, neuroendocrine, adipogenic, and cardiovascular systems. It is also an important factor in controlling the pubertal growth spurt, growth plate closure, and accretion of BMD of long bones.91011
Many mechanisms or theories attempting to explain these effects have been suggested. First, estrogen binds to ER in the pituitary somatotrope and activates the GH-IGF-I axis and it is believed to be a major factor for the pubertal growth spurt.12 Second, estrogen is involved in the stimulation of osteoblastogenesis, reduction of mature osteoblast apoptosis, suppression of osteoclastogenesis, and inhibition of osteoclastogenic cytokine production, and these actions are thought to be related with the control of BMD in long bones.1314 But, the effect of estrogen on the mechanism of growth plate closure is still unclear. There are only a few theories, including apoptosis, autophagy, hypoxia, and transdifferentiation of chondrocytes in the epiphyseal plate that maybe promoted by estrogen.15
If we could elucidate the precise process of growth plate closure and delay it without severe side effects, we may increase the final height of children more efficiently.
The most effects of estrogen are mediated by binding to ERs in the classical pathway. ER is a member of the nuclear receptor superfamily and functions as a ligand-inducible transcription factor4. There are two major subtypes, ERα (ESR1) and ERβ (ESR2) that are distributed in various tissues and function in distinct ways in several target tissues. ERα is mainly distributed in the uterus, breast, testis, hypothalamus, liver, heart, and skeletal muscles and ERβ is mainly distributed in the ovary and prostate. Both subtypes are present in bone, epididymis, thymus, adrenal, brain, and other parts of the body.161718
ER is also present in the epiphyseal plate, and there are 3 distinctive zones according to the distribution of different types of chondrocytes: the resting zone which is composed of stem cells of chondrocytes, the proliferative zone with increasing number of cells, and the hypertrophic zone composed of larger chondrocytes in the growth plate of long bones.19 ERα and ERβ are largely distributed in the resting and proliferative zones, although ERβ is slightly more prominent in the hypertrophic zone, which is involved in the transition of the chondrocyte to osteocyte in the epiphyseal plate.20
Börjesson et al.21 and Chagin et al.22 suggested that low E2 levels increase skeletal growth during the early sexual maturation and the pubertal growth spurt, whereas high E2 levels during late puberty result in growth plate fusion. If a higher E2 serum concentration is needed to activate ERβ than to activate ERα, it can be inferred that the activation of ERβ is essential for the growth plate fusion, and the activation of ERα is more important for the stimulation of the GH-IGF-I axis under low E2 levels. ERβ inhibits bone growth in mouse only when activated through increased estrogen serum levels, and the ERβ activation has the ability to induce growth plate fusion in old female mice. Therefore, we hypothesized that the selective inhibition of the ERβ activation might be a preferred method to delay the growth plate closure with lesser side effect on the pubertal growth spurt or BMD.
To test our hypothesis, we used synthetic ERα or ERβ agonist to stimulate each ER subtype selectively. It has been previously demonstrated that PPT is a potent ERα agonist, with a 400-fold preference for ERα over Erβ.2324 In contrast, DPN is a selective ERβ agonist with a 70-fold higher affinity to ERβ than to ERα.2526
In our study, after injections for 5 weeks, there were no significant differences in crown-rump length among 3 groups, but there was a significant decrease in the weight for the PPT group. This may mean that ERα mediates estrogen's anorexigenic effect or plays a role in suppressing white adipose tissue development in subcutaneous fat, and those effects are consistent with previous studies showing increased adipose tissue in an ERα knockout (KO) female mouse.2728
We also measured serum GH levels and pituitary Gh1 gene expression in each group. GH secretion was increased in PPT group and Gh1 expression was significantly increased in both PPT and DPN groups, but it was more prominent in the PPT group. This result suggested that the ERα activation is related with the stimulation of GH-IGF-I axis and the pubertal growth spurt, and is in agreement with a previous report by Avtanski et al.29
BMD was increased in PPT group without estrogenic effect after ovariectomy. Therefore, ERα stimulation is believed to be important for bone mineral deposition. This finding is consistent with the previous studies that the reduced BMD induced by estrogen deficiency by ovariectomy in animal models was recovered by E2 or ERα agonists. Khalid and Krum30 reported that signaling via ERα protects against ovariectomy-induced trabecular bone loss, and ERα activity can be modulated by ERβ in female bones. Lindberg et al.31 found that the ovariectomized wild or double ER knockout mice had the phenotype of increased cortical and trabecular bone dimension after E2 injection and it may mean the bone mineral deposition is mainly ERα mediated. Hertrampf et al.32 reported that the injections of the ERα-specific agonist (16α-LE2) increased BMD and serum bone formation markers but the ERβ-specific agonist (8β-VE2) did not in female rats.
In the studies about the growth and epiphyseal plate fusion of long bones, Chagin et al.33 reported that young adult ERβ−/− mice demonstrated an increased axial- and appendicular-skeletal growth, supporting that ERβ inhibits skeletal growth and has the capacity to mediate growth plate fusion. But, Iravani et al.34 reported that E2- and PPT-treated ovariectomized female mice had shorter tibia and femur bones, and their growth plate and hypertrophic zone height were decreased, which means the ERα is more important for growth plate fusion. Like these previous reports, some data are conflicting, which could be explained by strain differences or low numbers of animals.
In our study, neither ERα nor ERβ stimulation significantly affected the growth plate thicknesses. Perhaps, this is because the growth plates do not fuse directly after sexual maturation in rodents or the physiology of longitudinal bone growth is different between humans and rodents.
A few limitations of our study are the low number of individuals in the sample, the failure of getting the 24-hour growth hormone secretion profile and the fact that the physiological mechanism of growth plate senescence may be different between humans and rodents.
In conclusion, we evaluated the growth of the body length and weight, secretion of GH, acquisition of BMD, and fusion of epiphyseal plate after ERα and ERβ stimulation in ovariectomized female rats. Our study showed that the ERα activation is more important than the ERβ in the pubertal growth spurt with activation of the GH-IGH-I axis. ERα stimulation is also believed to be important for bone mineral deposition and prevention of weight gain. Therefore, the ERα agonists are thought to be effective for height growth, bone mineral deposition and weight loss.
But, the effects of activation of each ER subtype on bone growth are considered to be complex and mediated by multiple signaling pathways. Therefore, more studies are necessary to elucidate the mechanisms of action of each ER subtype in regulating the pubertal growth spurt and growth plate closure. In addition, in vitro studies on signaling pathways of each ER subtype and in vivo studies in other ERα or ERβ KO animal models are needed.
Go to :
 XML Download
XML Download