

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The primary brain tumor glioblastoma multiforme (GBM) is a rapid-growing and invasive tumor of glial origin with poor prognosis.1 The mean survival time of patients with GBM is approximately 1 year, whereas that with anaplastic astrocytoma is 2 to 3 years.2 Many determinants have been suggested as causative factors in the development of GBM, but the exact mechanism of the tumorigenesis is unknown.
Human cytomegalovirus (HCMV) has been centered on the association with glioblastoma, although some investigators reported the absence of HCMV in glioblastoma shown by high-coverage DNA sequencing3 and by immunohistochemistry and quantitative polymerase chain reaction.4 The level of HCMV infection in GBM is correlated with prognosis and may be involved in the pathogenesis of GBM. In fact, HCMV sequences and proteins have been found in most GBMs, but not in healthy brain.5 An immunotherapy clinical trial using dendritic cells pulsed with autologous glioblastoma lysate generated cytotoxic T lymphocytes (CTLs) specific to HCMV pp65.6 However, there are challenges for any pathogen or immune component to enter the central nervous system. Due to a very tight barrier structure, the blood–brain barrier, the brain is known as to be an immunologically privileged site, and it tolerates the antigenic challenges without any immune response.7 However, recent clinical trials with antibodies8 and experimental animal observations, such as experimental autoimmune encephalitis,9 have shown penetration of immune components into brain.
HCMV, a member of the Herpesviridae, is a double-stranded DNA virus with a genome of 230 kb, encoding for over 200 proteins. During productive infection, the replication cycle of HCMV is characterized by sequential expression of immediate early (IE), early (E), and late (L) gene regions. IE genes are expressed prior to the synthesis of the viral proteins immediately after an infection. After the expression of IE gene products, the E and L genes are successfully produced. HCMV infection is common, and the seropositivity of HCMV is 50%–100% worldwide.10
HCMV proteins are immunodominant antigens, and they evoke strong immune responses in humans after infection.11 Around 10% of peripheral CD8+ memory T lymphocytes are reactive to HCMV proteins. The number increases with age, eventually comprising over 45% of the total CD8+ T lymphocytes.12
HCMV has neurotrophic properties. A fetus can be infected with HCMV through transplacental transmission from the mother, and it results in damage to the brain such as microcephaly, deafness, and mental retardation. A recent study of the congenital infection with mouse cytomegalovirus in an animal model has broadened the understanding of central nervous system infection.13 The recruitment of immune cells and the expression of pro-inflammatory cytokines are involved in brain impairment. The perivascular infiltration of mononuclear cells was seen in GBM and is correlated with prognosis.14 HCMV IE-1 was also detected in 93%–100% of GBM.5 Recent our report suggested that HCMV inhibits local immune responses via direct killing of immunocytes near HCMV-infected cells through reactive oxygen species-induced parthanatos by HCMV-associated insoluble substances.15
To examine the correlation of viral infection with tumor expansion, we prepared CTLs specific to HCMV IE-1 and demonstrated that they lysed HCMV-infected astrocytoma cells in a dose-dependent manner, but they showed decreased functioning in the presence of a small number of effector cells.
Here, we sought to verify a role of HCMV-infected astrocytoma cells in impairing the activity of CTLs specific to HCMV IE-1.
METHODS
Cells and virus
U373MG cells (ATCC HTB-17: male) constitutively expressing HCMV IE-1, UMG1-2,16 were used as stimulating cells and target cells for cytotoxic T cells (CTL) to HCMV IE-1. U373MG transfected with empty LNCX vector, were used as control cells.17 Jurkat cells (ATCC TIB-152: male) were used as control T cell source. The cells were cultured in Dulbecco's modified Eagles medium (DMEM; GIBCO, Grand Island, NY, USA) with 10% fetal bovine serum (FBS; GIBCO), 100 U/mL penicillin and 100 U/mL streptomycin (Life Technologies, Carlsbad, CA, USA) in a 37°C incubator with 5% CO2. HCMV Towne strain (ATCC VR-977) was used to infect cells at a multiplicity of infection of 4 throughout the experiment.
Peripheral blood mononuclear cells (PBMCs) of 6 healthy donors were collected at the interface on Ficoll-Paque (GE Healthcare, Little Chalfront, UK) after centrifugation at 700 g for 30 minutes. CD8+ T cells were isolated with CD8+ T cell isolation kit (Miltenyi Biotech, Surrey, UK) and MACS LS column (Miltenyi Biotech) according to the manufacturer's manual. PBMCs and CD8+ T cells were cultured in RPMI1640 (Thermo Scientific, Waltham, MA, USA) containing 10% FBS in 5% CO2 atmosphere at 37°C.
Antibodies and reagents
Monoclonal anti-HCMV IE-1 (clone 6IE1) and fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG (Life Technologies) were used to detect HCMV IE-1 antigen. Phycoerythrin (PE)-conjugated anti-HLA-A0201 (BD Biosciences, San Jose, CA, USA) was used to detect the expression of HLA antigen on the cells.
HCMV IE-1 overlapping peptides with 11 amino acids from IE-1 protein of HCMV AD169 (Miltenyi Biotech) were used to expand CTLs specific to HCMV IE-1. All reagents were purchased from Sigma Chemical (St. Louise, MO, USA) unless otherwise stated.
Expansion of CTLs specific to HCMV IE-1
PBMCs or CD8+ T cells from donors were added onto UMG1-2 cells, cultured for 48 hours with the intermittent gentle shaking. Cells in the supernatant were harvested, washed with media containing 10% FBS, and used as HCMV IE-1-specific CTLs.
Flow cytometry analysis
To analyze the surface expression of major histocompatibility complex (MHC) I, human leukocyte antigen (HLA)-A*0201, on cells, cells were collected, washed in Flow Cytometry Staining (FACS) buffer (phosphate-buffered saline with 0.5% bovine serum albumin, 0.01% NaN3), and reacted with PE-conjugated anti-HLA-A*0201 on ice for 30 minutes. After washing with FACS buffer, fluorescent signals were analyzed with FACS LSR. In case of the detection of HCMV IE-1, the collected cells were permeabilized with 70% ethanol on ice for 30 minutes, and reacted with anti-HCMV IE-1 antibody for 30 minutes, followed by FITC-conjugated with goat anti-mouse IgG for 30 minutes.
Western blot analysis
Western blot analysis of HCMV IE-1 was performed at different time points after HCMV infection. Protein concentration in cell lysates was determined with BCA protein assay kit (Thermo Scientific) as described in the manufacturer's instruction manual. Samples with 100 μg/lane were subjected on a denaturing sodium dodecyl sulfate 10% polyacrylamide gel under reducing conditions by electrophoresis. Separated proteins were electrophoretically transferred to polyvinylidene difluoride (PVDF; Millipore, Darmstadt, Germany) membranes. After the appropriately diluted anti-HCMV IE-1 antibody was reacted for 24 hours at 4°C, horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (Calbiochem, San Diego, CA, USA) was used as a secondary antibody. Signal was developed using an enhanced chemiluminescence reagent (Thermo Scientific) as described in the manufacturer's instruction manual, and visualization was achieved by LAS-4000 (Fujifilm, Tokyo, Japan).
Assays for cytotoxic activity
The cytotoxic activity of CTLs was assayed by a viable cell counting of target cells. Target cells pre-stained with cell proliferation dye 670 (eBioscience, San Diego, CA, USA) were incubated with effector cells for 24 hours, harvested and analyzed with flow cytometry. The total number of target cells without the effector cells was used as a control. Cell death rate was calculated with the formula of (number of control − number of target cells after the incubation with effectors)/number of control × 100. All experiments were repeated three times independently and the result was expressed as mean ± standard error.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
Cells undergoing death was determined by flow cytometry using TUNEL staining methods using APO-BRDU™ Kit (Millipore) according to the manufacturer’s instruction. Approximately 1 × 106 cells were fixed with 70% ethanol and stored overnight at −20°C. The cells were washed twice with washing buffer, followed by incubation for at least 1 hour with staining solution at room temperature. Staining solution contained 1 mg/mL RNase A and TdT reaction reagent and antibody solution containing BrdU-FITC. DNA content or DNA fragmentation was measured in cells with a FACS Fortessa (BD Biosciences). Data of cell cycle were analyzed using FlowJo cytometry analysis software (Tree Star, Ashland, OR, USA). The cell death experiments were repeated three times and the results were expressed as mean ± standard error.
Quantitation of HCMV
The copy number of HCMV was assayed by real-time polymerase chain reaction (PCR). Sample DNA was prepared with Media kit (Qiagen, Valencia, CA, USA) from culture supernatant according to the manufacturer's recommendation. Real-time PCR reactions were performed using a Taqman MasterMix (Applied Biosystems, Foster City, CA, USA) with 10 pmol HCMV US17 primer pairs, (5′- GCGTGCTTTTTAGCCTCTGCA-3′) and (5′-AAAAGTTTGTGCCCCAA CGGTA-3′), 10 pmol probe, FAM-5′-TGATCGGCGTTATCG CGTTCTTGATC-3′-TAMRA, and sample DNA in a 20 μL reaction with an ABI PRISM 7900 sequence detection system (Applied Biosystems). Primers and probes were synthesized commercially at Bioneer (Daejeon, Korea). Reactions were performed under standard universal reaction conditions; hot start cycle of 10 minutes at 95°C, and 40 cycles of denaturation for 10 seconds at 95°C and annealing and extension for 60 seconds at 60°C. Standard curve for the quantitation of HCMV was obtained with the serially diluted samples of the known amount of HCMV.
RESULTS
Characteristics of cells used in experiments
The phenotype of the HLA-A on cells was determined by FACS analysis. U373MG, UMG 1-2, and PBMCs from three donors, donors A1, A2 and A3, expressed HLA-A*0201, but PBMCs from three other donors, donors C1, C2 and C3, did not (Supplementary Fig. 1).
Expansion of HCMV IE-1-specific CTLs
The presence of HCMV IE-1-specific CTLs in PBMCs from the donors was identified. When PBMCs from HLA-A*0201(+) donor A1 were stimulated with overlapping peptides of HCMV IE-1, CD8+ T lymphocytes secreting interferon (IFN)-γ were detected with 1.99%. This secretion was not detected prior to stimulation. These results demonstrate the presence of HCMV IE-1-specific CTLs in donor A1 (Supplementary Fig. 2A).
For the effective and physiological production of HCMV IE-1-specific CTLs, CD8+ T lymphocytes were purified from PBMCs, and the cell line UMG1-2 was used as a stimulator. The purity of CD8+ T lymphocytes was 71.8% (Supplementary Fig. 2B). Expression of HCMV IE-1 was detected in UMG1-2 cells, but not in the U373MG cells used as a negative control (Fig. 1A). Expression of HCMV IE-1 in U373MG cells after HCMV infection was determined by Western blot analysis and FACS. Both analytic methods showed that HCMV IE-1 increased in U373MG cells infected with HCMV from day 1, reached at maximum 2–3 days post-infection, and declined thereafter. U373MG cells were used as a negative control. CTLs were expanded by the stimulation of purified CD8+ T lymphocytes from the HLA-A*0201(+) donor with UMG1-2 cells for 2 days. Increased morphological changes and decreased confluency in UMG1-2 cells compared with U373MG cells were observed under a microscope when each cell was co-cultured with an increasing number of CTLs (Fig. 1B). Confluency of cells was decreased only when CTLs from HLA-matched donor were treated to UMG1-2 cells. The death rates of the UMG1-2 cells were 19.0% ± 26.9% at E:T = 1:1 and 73.9% ± 8.2% at E:T = 3:1 after incubation with CTLs generated as described above (Fig. 1C). In the case of CTLs from HLA-A*0201-negative donor, the death rates of UMG1-2 were 4.6% ± 5.9% and 22.4% ± 4.2% at E:T = 1:1 and 3:1, respectively. When the target cell was U373MG, the cell death rate was below 15.3% in all assays (Fig. 1C).
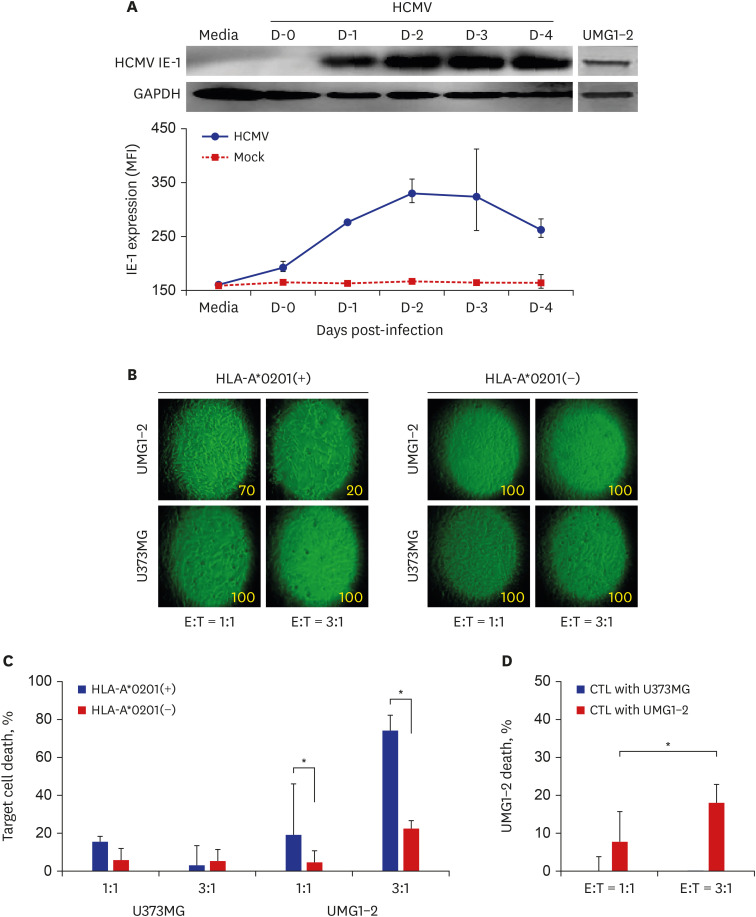
Fig. 1
Expansion of HCMV IE-1-specific CTLs from CD8+ T lymphocytes. (A) Expression profile of HCMV IE-1 determined by western blot analysis and FACS. HCMV IE-1 was detected in HCMV-infected U373MG cells on the indicated time points. UMG1-2 cells were used as positive control of HCMV IE-1 expression. (B) Representative photographs of UMG1-2 and U373MG cells co-cultured with the prepared CTLs. The morphological changes increased and cell confluency decreased in UMG1-2 cells by the adding of the increasing number of CTLs, which were expanded by the stimulation of CD8+ T lymphocytes from HLA-A*0201(+) donor with UMG1-2 cells. Number in each photograph means the estimated cell confluency (%) observed under microscope. (C) The death rate of the target cells by CTLs expanded by stimulation with UMG1-2. The death of UMG1-2 cells increased after the culture with the increasing number of CTLs, which were expanded by the stimulation of CD8+ T lymphocytes from HLA-A*0201(+) donor with UMG1-2 cells. Significant cell death of UNG1-2 was found only in CTLs from HLA-A*0201(+) donor. (D) The death rate of the target cells, UMG1-2, by CTLs expanded by stimulation with U373MG (CTLs with U373MG) or UMG1-2 (CTLs with UMG1-2). The death of UMG1-2 cells increased after the culture with the increasing number of CTLs, which were expanded by the stimulation of CD8+ T lymphocytes from HLA-A*0201(+) donor with UMG1-2 cells (CTLs with UMG1-2), but not with U373MG (CTLs with U373MG).
HCMV = human cytomegalovirus, IE = immediate early, CTLs = cytotoxic T lymphocytes, FACS = Flow Cytometry Staining, HLA=human leukocyte antigen.
*P < 0.05, χ2 test.
![]()
For the verification of the specificity of CTLs to HCMV IE1, CTLs were expanded by the stimulation of purified CD8+ T lymphocytes from the HLA-A*0201(+) donor with U373MG or UMG1-2 cells for 2 days. The death rates of the UMG1-2 cells were −0.6% ± 4.3% and 7.8% ± 4.1% at E:T = 1:1, respectively, and −6.2% ± 7.9% and 18.0% ± 4.9% at E:T = 3:1, respectively, after incubation with CTLs generated with U373MG or UMG1-2 as described above (Fig. 1D).
Death of CTLs by HCMV-infected U373MG cells
Jurkat cells were added to HCMV-infected U373MG cells on the indicated day and incubated for 4 hours. The harvested Jurkat cells were stained with Annexin V, and cell death was analyzed using flow cytometry. From day 0 to day 1 post-infection, cell death was observed in the Jurkat cells. However, on day 2, post-infection cell death increased to 30% and then rapidly increased up to 80%–90% thereafter. No cell death was seen in the Jurkat cells in the control experiment (Fig. 2A).
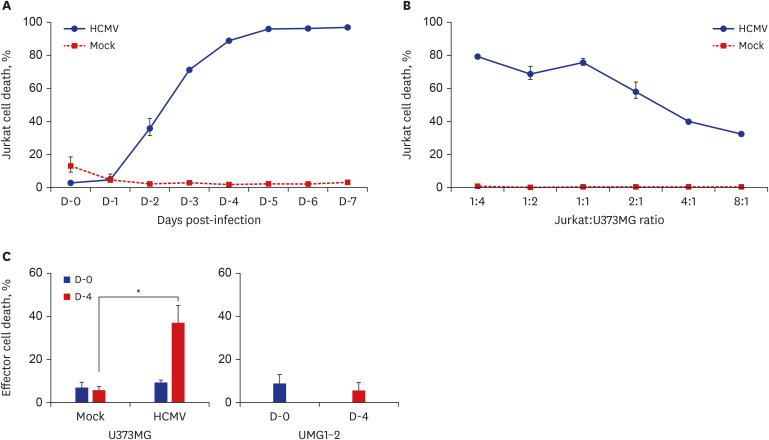
Fig. 2
The death of CTLs by HCMV-infected U373MG. (A) The death of Jurkat cells by HCMV-infected U373MG cells depending on the stage of infection. Jurkat cells were added into HCMV-infected U373MG cells on the indicated day, incubated for 4 hours, and the death of the harvested Jurkat cells were determined by annexin V staining assay. Cell death increased from day 2 post-infection and reached at maximum at day 4 post-infection. (B) The death of Jurkat cells by HCMV-infected U373MG depending on the adding number of Jurkat cells. Jurkat cells were added into HCMV-infected U373MG cells on day 4 post-infection, co-cultured for 24 hours, and the death of the harvested Jurkat cells were determined by TUNEL assay. The death rate of Jurkat cells maintained at a constant level in the co-cultures of 1:4 to 1:1, but decreased proportionally in the cultures of 2:1 to 8:1. (C) The death of CTLs by HCMV-infected U373MG cells. CTLs were added into HCMV-infected U373MG cells at day 0 and 4 post-infection, co-cultured for 24 hours, and the death of the harvested CTLs were determined by TUNEL assay. Increased death rate of CTLs was found in the co-cultures with HCMV-infected U373MG cells, not in the co-cultures with U373MG or UMG1-2 cells.
CTLs = cytotoxic T lymphocytes, HCMV = human cytomegalovirus, TUNEL = terminal deoxynucleotidyl transferase dUTP nick end labeling.
*P < 0.05, χ2 test.
![]()
To determine the death-inducing capacity of HCMV-infected U373MG cells to effector cells, a cytotoxic assay was performed using a number of ratios of Jurkat cells to HCMV-infected U373MG cells. Variable numbers of Jurkat cells were added to U373MG cells infected with HCMV for 4 days. After co-culture for 24 hours, a cytotoxic assay was performed using the TUNEL assay. The cell death rates of Jurkat cells when added to HCMV-infected U373MG cells at ratios of 1:4, 1:2, and 1:1 were 79.2% ± 4.7%, 68.7% ± 2.5%, and 75.7% ± 5.7%, respectively, but using ratios of 2:1, 4:1, and 8:1, the cell death rates were 58.1% ± 1.5%, 40.1% ± 0.2%, and 32.5% ± 1.2%, respectively (Fig. 2B). These results suggest that as more effector cells are present in the culture of HCMV-infected U373MG cells, the death rate of the effector cells decreases due to the limited death-induction capacity of HCMV-infected U373MG cells.
Two time points, day 0 and day 4 post-infection, were selected to assay the death of CTL by co-culture with HCMV-infected U373MG cells to observe the effect of the productive infection of HCMV on the function of CTLs. When CTLs generated from CD8+ T lymphocytes from donor A1 were added to the culture, the cell death rates of the CTLs were 9.4% ± 1.0% at day 0 and 37.1% ± 7.9% at day 4 post-infection. In the control cultures using a mock infection and UMG1-2 cells, the death rate was below 8.8% in all assays (Fig. 2C). HCMV was quantitated using the culture supernatant from HCMV-infected U373MG cells at day 4 post-infection by real-time PCR. The mean amount of HCMV was 4.5 × 104 PFUs/mL.
Changes in MHC I antigen on the cells due to HCMV infection
The expression level of MHC class I on U373MG cells after HCMV infection was monitored with anti-HLA-A*0201 antibody using FACS analysis. At day 0 post-infection, the level decreased to 94.1%, and it was maintained at 87.3%–97.4% until day 3 post-infection. It then decreased to 61.3%–67.6% at days 4–5 post-infection and to 35.1% at day 6 post-infection. It did not change significantly in the control mock-infected U373MG cells throughout the experimental period (Fig. 3A).
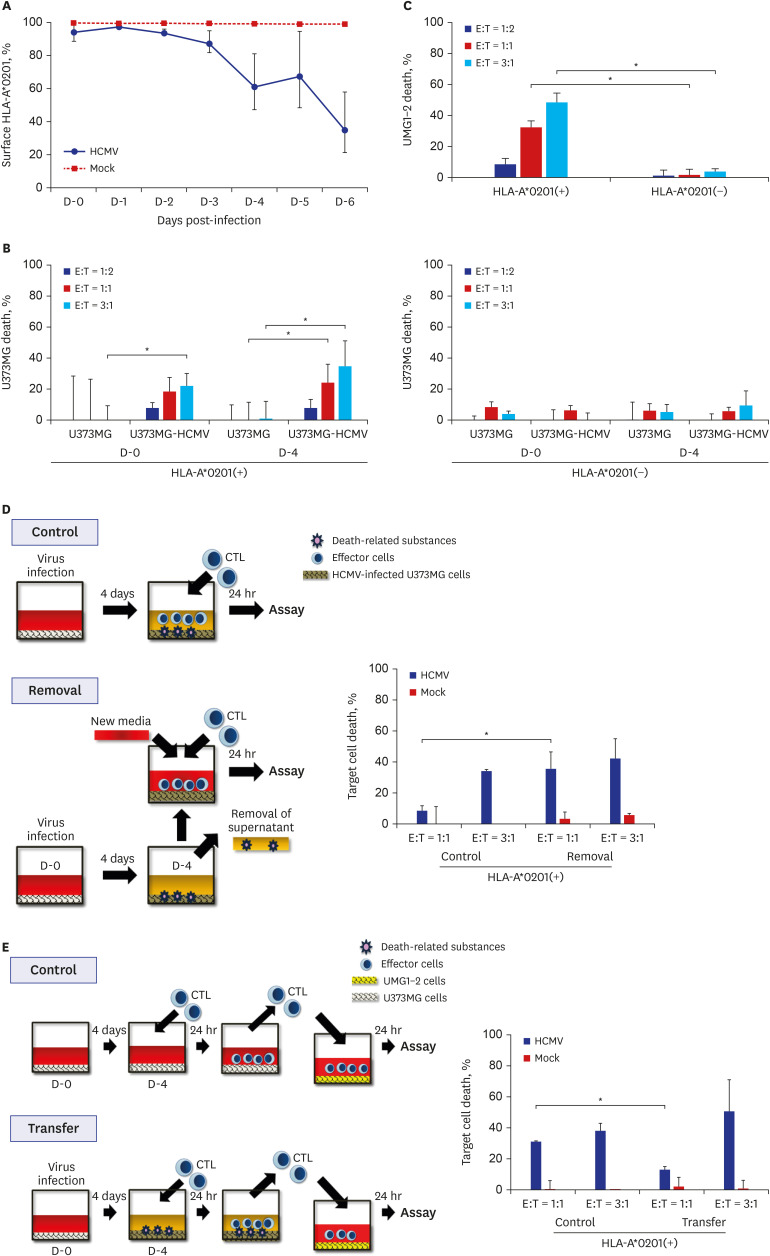
Fig. 3
The death of HCMV-infected U373MG cells by CTLs. (A) Changes of the MHC I expression on U373MG cells infected with HCMV. Expression of MHC I on U373MG infected with HCMV was assayed by FACS analysis after the staining with the anti-HLA-A*0201 antibody in the indicated time points. MHC I expressed on HCMV-infected U373MG cells decreased significantly 4 days after infection. (B) The death rate of HCMV-infected U373MG cells by CTLs, which were expanded by the stimulation of CD8+ T lymphocytes from HLA-A*0201(+) or (−) donor with UMG1-2 cells. The data were expressed as mean ± standard deviation from three independent assays. The cell death rate increased depending on HLA restriction and CTL dose. (C) The death rate of UMG1-2 cells by CTLs, which were expanded by the stimulation of CD8+ T lymphocytes from HLA-A*0201(+) or (−) donor with UMG1-2 cells. The data were expressed as mean ± standard deviation from three independent assays. The cell death rate increased depending on HLA restriction and CTL dose. (D) Effect of the removal of the culture supernatant from HCMV-infected U373MG cells on the killing of target cells, HCMV-infected U373MG cells, by CTLs. Removal of the culture supernatant from HCMV-infected U373MG enhanced the death of the target cells at E:T = 1:1, but not at E:T = 3:1. The results were taken from two donors, and the data were expressed as mean ± standard deviation from three independent assays. (E) Effect of the transfer of CTLs from 24-hour co-culture of HCMV-infected U373MG cells and CTLs on the killing of the target cells, UMG1-2 cells. Transfer of them to UMG1-2 cells decreased the death of the target cells at E:T = 1:1, but not at E:T = 3:1. The results were taken from two donors, and the data were expressed as mean ± standard deviation from three independent assays.
HCMV = human cytomegalovirus, CTLs = cytotoxic T lymphocytes, MHC = major histocompatibility complex, HLA = human leukocyte antigen, E:T = effector:target.
*P < 0.05, χ2 test.
![]()
Cell death of HCMV-infected U373MG cells by CTLs
The cell death rate in HCMV-infected U373MG cells was measured after the treatment of CTLs generated from the stimulation of purified CD8+ T lymphocytes of HLA-A*0201(+) or (–) donors with UMG1-2 cells. The cell death rate by CTLs increased from 18.7% ± 8.8% to 24.3% ± 11.7% at day 0 and to 22.1% ± 8.1% to 34.7% ± 16.3% at 4 days post-infection by increasing the E:T ratio from 1:1 to 3:1 in HLA-A*0201(+) donors. It was below 9.4% in all assays when the CTLs from HLA-A*0201(–) donors were used (Fig. 3B).
The cell death rates in UMG1-2 by CTLs generated as above were 8.7% ± 3.7%, 32.3% ± 4.3%, and 48.2% ± 6.2% at E:T = 1:2, 1:1, and 3:1, respectively. When CTLs were generated from the HLA-A* 0201(–) donor, they were under 3.8% at all E:T ratios (Fig. 3C). These results demonstrated that the generated CTLs could kill target cells depending on the HLA restriction and the amount of CTLs.
When the culture supernatant from HCMV-infected U373MG cells were removed and CTLs were added to HCMV-infected U373MG cells, the target death increased from 8.4% ± 8.4% to 40.8% ± 14.3% at E:T = 1:1 (P < 0.05), but not at E:T = 3:1 (41.5%–51.2%, P > 0.05) (Fig. 3D). When CTLs were collected from 24-hour co-culture of HCMV-infected U373MG cells and CTLs, and they were transferred to UMG1-2 and cultured for a further 24 hours, the cell death rate of the target cells, UMG1-2, decreased from 31.1% ± 0.7% to 13.0% ± 2.0% at E:T = 1:1 (P < 0.05), but increased from 38.0% ± 5.0% to 50.9% ± 20.4% at E:T = 3:1 (P > 0.05) (Fig. 3E).
DISCUSSION
We demonstrated that CTLs prepared in this study could lyse the target cells, HCMV-infected astrocytoma cells, depending on the number of CTLs present, but they had limited lytic activity on the target cells in the case of a relatively small number of CTLs in the late stages of infection.
HCMV pp65 and IE-1 have been found to be major targets of CTL response.1819 HCMV pp65 may be more suitable than HCMV IE-1 for candidate molecules of a vaccine or immunotherapy because both CD4+ and CD8+ T lymphocytes reactive to HCMV pp65 were detected in many cases, whereas only CD8+ T lymphocytes reactive to HCMV IE-1 were detected.20 However, the situation is complicated in the immune response for GBM. Once the HCMV genome is introduced into the cell, the major immediate early promoter (MIEP) of HCMV can be activated, IE genes are transcribed, and IE proteins are produced in almost all cells. The susceptibility and permissiveness to HCMV infections vary in different cell lines derived from the same tissue origin, and even from cell to cell of the same tissue. HCMV IE-1 was detected in all GBM.21 HCMV pp65 and HCMV IE-1 were present in fresh isolated GBM, but HCMV pp65 was less ubiquitous than HCMV IE-1.5 The HCMV late antigen, for example pp65, cannot be produced in the cell if viral replication has not occurred for any reason, whereas HCMV IE-1 can be produced in the cell after HCMV infection regardless of viral replication. Therefore, we chose HCMV IE-1 as a target molecule to produce the specific CTLs for the study of reactivity to astrocytoma cells infected with HCMV. We showed that CTLs to HCMV IE-1 could react to cells expressing HCMV IE-1 throughout the whole period of viral infection, from the early (Fig. 3B, D-0) to the late stages (Fig. 3B, D-4).
The presence of CTLs reactive to HCMV IE-1 was demonstrated by stimulation of PBMCs from the HLA-A*0201(+) donor with overlapping peptides of HCMV IE-1 and probing of IFN-γ-secreting cells (Supplementary Fig. 2A), which result is comparable to other's report with 2%–3% activated cells.22 For the effective production of CTLs to HCMV IE-1, UMG1-2 cells were used as a stimulator. When the HLA phenotype of the donors was HLA-A*0201(+), the CTLs expanded with UMG1-2 had specific lytic activity to UMG1-2 cells with CTL dose dependence, but not to U373MG cells, while the CTLs expanded with U373MG did not show any lytic activity to UMG1-2 cells (Fig. 1B-D). These results suggest that CTLs specific to HCMV IE-1 rather than U373MG cells themselves were predominantly expanded with the stimulation of UMG1-2, and react to target cells effectively.
HCMV has many strategies for immune evasion, such as the decreased expression of MHC I molecules on the infected cells.23 But effective CTLs reactive to HCMV-infected cells can be produced in healthy individuals. This means that the immune system of the host overcomes the evasion attempt of HCMV. To date, no reported study has examined the death of effector cells in CTL assays. We have demonstrated that when the effector cells in cultures of CTLs and HCMV-infected U373MG cells were incubated for 24 hours the cell death increased up to 37.1% at day 4 post-infection, whereas in the cultures of CTLs and U373MG cells, the death rate of the effector cells was below 8.8% (Fig. 2C). This result suggests that HCMV-infected U373MG cells at day 4 post-infection produced factors that impaired the function of CTLs. Reactive oxygen species-induced parthanatos of immunocytes nearby HCMV-infection sites by human cytomegalovirus-associated substance15 would be one of many candidates of mechanisms. The further detailed mechanism of the death of effector cells by HCMV infection should be elucidated in the future studies.
Many tumors express variant MHC molecules,2425 and it is feasible to use the antibody to the exact polymorphic MHC epitope as a probe. We monitored the expression of MHC I molecules on U373MG cells after HCMV infection with anti-HLA-A*0201 antibody instead of antibody to common MHC I or b2 microglobulin. MHC I molecules remained above 80% until day 3 post-infection and decreased to 40%–60% thereafter (Fig. 3A). This decreased level of MHC I on U373MG cells after HCMV infection contradicts by another publication.26 The difference may derive from the use of different antibodies. The death rate of HCMV-infected U373MG cells by CTLs rose to 41.9% at E:T = 3:1 with CTL dose-dependence at day 4 post-infection, although the expression of MHC I on HCMV-infected U373MG cells decreased to 61.3%. These data suggest that the immune reaction to tumor cells infected with HCMV might be different from that to the normal cells infected with HCMV.
We were unable to recruit a HCMV-seronegative donor because almost all individuals in Korea are seropositive.27
We removed the supernatant from the cultures of HCMV-infected U373MG cells before adding CTLs to see the effect of the culture supernatant on the death of CTLs. As expected from the simulation of the death of the effector in Fig. 2C, the cell death rate of the target cells increased from 8.4% to 40.8% at E:T = 1:1 after the removal of the culture supernatant, but did not change at E:T = 3:1 (Fig. 3D). Similarly, with the transfer of the cells to UMG1-2 after the co-culture of CTLs and HCMV-infected U373MG for 24 hours, the cell death rate of CTLs decreased from 31.0% to 13.0% at E:T = 1:1, but not at E:T = 3:1 (Fig. 3E). These results implied that CTLs specific to HCMV IE-1 could lose their function near HCMV-infected U373MG cells when the number of CTLs was relatively low. The capacity of HCMV-infected U373MG cells to induce the death of nearby target cells seems to be limited. When the relatively high number of Jurkat cells, which may be regarded as equivalent to effector cells, the death rate of Jurkat cells by HCMV-infected U373MG cells decreased compared with the experiment with the low number of Jurkat cells (Fig. 2B). Therefor it is reasonable to use the high number of CTLs specific to HCMV for the treatment of GBM infected with HCMV.
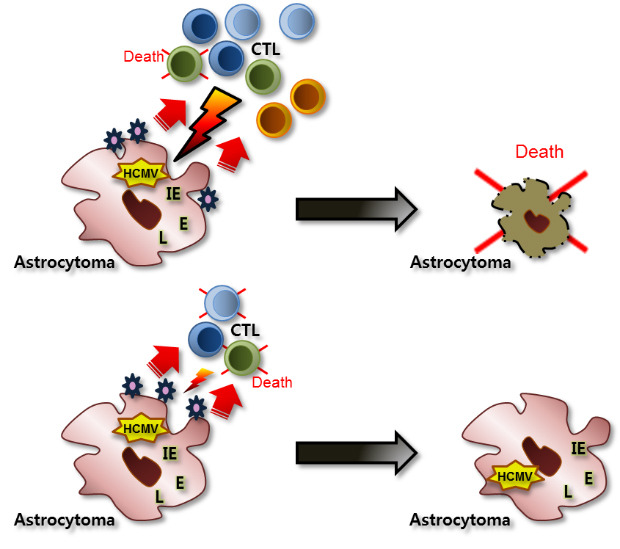
We have summarized the experimental results and presented models of CTL activities according to different microenvironments of HCMV infection and permissiveness of cells in astrocytes (Fig. 4). In the case of non-productive HCMV infection, for example, the early stage of HCMV infection or the viral entrance into cells without viral replication, CTLs specific to HCMV IE can lyse HCMV-infected astrocytes effectively. However, when HCMV replicates productively in astrocytes, the lytic capacity of CTLs specific to HCMV IE is dependent on the relative number of CTLs near HCMV-infected astrocytes. With the relatively small number of CTLs, the portion that are dead and the remaining CTLs have limited lytic activities relative to HCMV-infected astrocytes. However, a relatively large number of CTLs can overcome the death of the small fraction, revealing the effective lytic activities to HCMV-infected astrocytes.
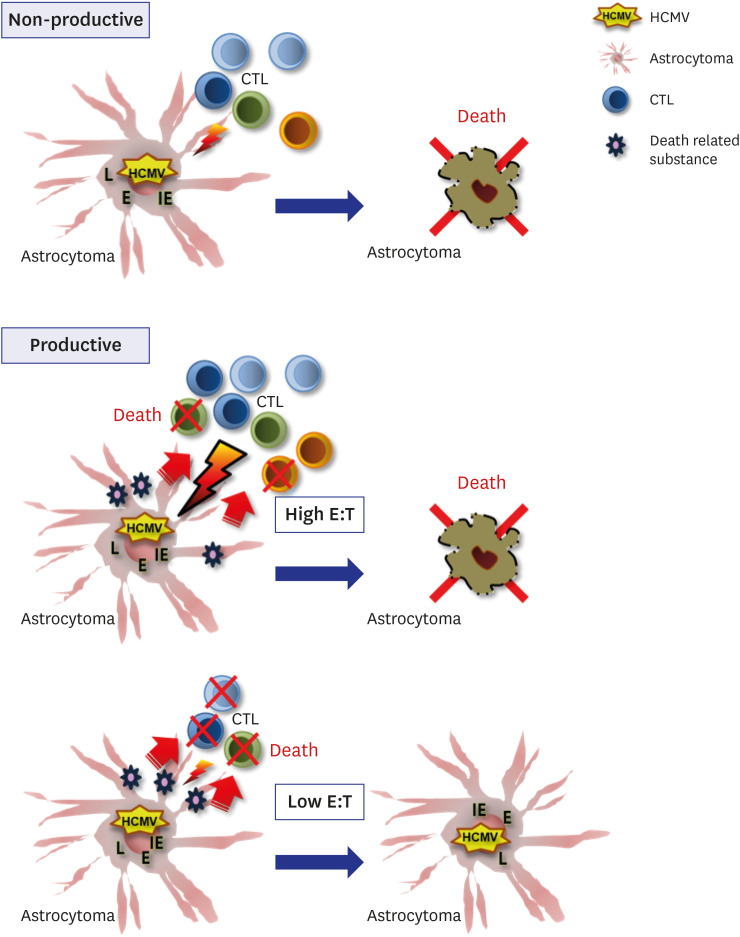
Fig. 4
Schematic presentation of the reactivity or the impairment of CTLs specific to HCMV by HCMV-infected astrocytoma cells according to the non-productive and productive microenvironment of HCMV infection.
CTLs = cytotoxic T lymphocytes, HCMV = human cytomegalovirus, L = late, E = early, IE = immediate early, E:T = effector:target.
![]()
In conclusion, HCMV-infected U373MG cells impair the activity of CTLs specific to HCMV when a relatively low number of the effector cells are present. These results suggest that HCMV infection impairs CTL function near HCMV-infected cells and facilitates glioblastoma growth unchecked by CTLs in a special micro-environment.
 XML Download
XML Download