

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The global spread of carbapenemase-producing (CP) Klebsiella pneumoniae (KPC) has become a major threat to human health [1]. Approximately 700,000 people die annually from antimicrobial-resistant infections, and this number is projected to surpass 10 million per year by 2050 [2]. This global phenomenon has been mainly associated with the dissemination of high-risk clones. The well-known KPC clones include the Kp sequence type (ST) 258 [3] and its related variants, which belong to clonal group 258 (CG258) [4]. However, in recent years, new drug-resistant lineages have emerged globally [5]. Among them, KPC-2-producing K. pneumoniae (KPC-Kp) ST307 is one of the most clinically relevant strains because its emergence has been recognized in several countries in the last five years [6, 7]. A study on non-CG258 clones of carbapenem-resistant K. pneumoniae (CR-Kp) strains spanning two yrs in Colombia showed that 62.2% of the strains were derived from ST307. Patients infected with KPC-Kp ST307 showed a high mortality rate (>50%) and had longer hospital stays than patients infected with other clones, indicating that this lineage encodes additional factors that contribute to its virulence [8]. In addition, the ST307 genome encodes various pathogenic genetic characteristics. The highly mobile Tn3-based transposon Tn4401 facilitates dissemination of the blaKPC gene [9]. The dissemination of blaKPC is derived by the clonal spread of bacterial hosts harboring the gene or by horizontal transfer of transposons and plasmids carrying the gene [10,11]. IncX3-type plasmid is prevalent in K. pneumoniae carrying the blaKPC gene [12].
As Kp ST307 is a relatively new strain, current knowledge on the genetic characteristics that facilitate its antimicrobial resistance and spread is limited. Thus, to enhance our understanding of Kp ST307 and facilitate the development of an effective therapy against infections caused by this strain, we investigated the genetic characteristics of KPC-Kp ST307 that contribute to its growth and spread, its antimicrobial resistance, and transfer among different Enterobacteriaceae strains. Additionally, we performed a whole-genome sequence analysis of KPC-Kp ST307 and other bacterial strains isolated from a single patient and investigated the horizontal transfer of plasmids containing the promiscuous blaKPC gene across intestinal bacterial strains of the patient. The whole-genome sequences were then compared and analyzed using minimum inhibitory concentration (MIC) analysis.
Go to : 
MATERIALS AND METHODS
Sources of KPC-producing Enterobacteriaceae isolates
The Institutional Review Board of BHS Hanseo Hospital, Busan, Korea, approved this prospective study (approval number: CTS-19-003). An 87-year-old woman with a history of primary knee arthropathy in both knees was admitted to a general hospital in Busan in August 2018 for arthritis treatment. CR Enterobacteriaceae were not detected in a rectal swab at admission. However, after one month, a urine culture test performed because of fever, confirmed KPC-2-producing Kp (CPKp1825). Additionally, Klebsiella aerogenes (CPEa1826) and Escherichia coli (CPEc1827) were detected from rectal swabs at 23 and 26 days, respectively, following the detection of the CPKp1825 strain in the urine culture. After the isolation of CPKp1825, vancomycin-resistant Enterococcus faecium was isolated from urine cultures repeated once every three days for two months, and the stool tested positive for Clostridioides difficile toxin.
Bacterial isolates and antimicrobial susceptibility testing
We identified clinical isolates using standard microbiological procedures and the identity of all isolates was confirmed by 16S rRNA gene sequencing [13]. Antimicrobial susceptibilities were determined by VITEK 2 AST N224 cards (bioMérieux Vitek, Hazelwood, MO, USA); susceptibility against 12 antimicrobial agents —ampicillin, aztreonam, cefotaxime, ceftazidime, cefoxitin, ciprofloxacin, amikacin, gentamicin, imipenem, meropenem, ertapenem, and colistin— was evaluated by the disk diffusion method on Mueller-Hinton (MH) agar (Becton Dickinson, Franklin Lakes, NJ, USA), following the Clinical and Laboratory Standards Institute (CLSI) guidelines [14]. Colistin susceptibility was determined by the broth microdilution method with freshly prepared cation-adjusted MH broth, following the CLSI guidelines [14]. Tigecycline susceptibility was confirmed by an E-test (bioMérieux, Marcy-l’Etoile, France). For the modified carbapenem inactivation method (mCIM) test, the bacteria were incubated with a meropenem disk in tryptic soy infusion (TSI; Difco Laboratories, Detroit, MI, USA) broth. For the EDTA-modified carbapenem inactivation method (eCIM) test, EDTA was added to the broth to chelate metal ions necessary for the metallo-β-lactamase function. After incubation, the disks were removed and placed on a lawn of susceptible E. coli to determine whether the test bacteria hydrolyze meropenem. Zone diameters were measured and interpreted following the CLSI guidelines [14].
Detection of resistance genes
We screened Kp strains for genes encoding extended-spectrum β-lactamases (ESBLs) (blaCTX-M-1-, blaCTX-M-9-, blaTEM-, and blaSHV-type), carbapenemases (blaIMP-1-type, blaVIM-2-type, blaNDM, blaKPC, blaKPC-2, blaGES, and blaOXA-48-like) [15], and outer membrane proteins (ompK35 and ompK36) using PCR (Verity 96-Well; Applied Biosystems, Eatate, Singapore) [16]. The PCR products were sequenced using a 3730xl automatic sequencer (Applied Biosystems, Weiterstadt, Germany), and the nucleotide sequences were compared using the Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi) (Supplemental Data Table S1).
Multilocus sequence typing (MLST)
We performed MLST PCR and sequencing for seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB for Kp [17], and adk, fumC, gyrB, icd, mdh, purA, and recA for E. coli) [18]. Nucleotide sequences obtained for both DNA strands were compared with sequences in the MLST database for each species (http://bigsdb.web.pasteur.fr/klebsiella for Kp and http://mlst.warwick.ac.uk/mlst/dbs/Ecoli for E. coli), to determine allelic numbers and STs.
Bacterial conjugation
Plasmids carrying the blaKPC gene in the CPKp1825, CPEa1826, and CPEc1827 strains were used as donors, and sodium azide-resistant E. coli J53 was used as the recipient [19]. For the standard agar mating method, equal amounts of overnight donor and recipient cultures were mixed and immediately spread on brain heart infusion (BHI; MBcell, Los Angeles, CA, USA) agar. After overnight mating at 37°C, bacterial cells were recovered and plated on MacConkey agar containing sodium azide (100 μg/mL) and imipenem (0.5 μg/mL) to select transconjugants.
Curing test
We performed KPC gene-carrying plasmid curing using the temperature-mediated plasmid emission method. CR Enterobacteriaceae strains were subjected to elevated temperature-mediated plasmid elimination through sequential passages in BHI (MBcell) broth twice per day for two weeks. Thereafter, the cultures were diluted and plated on TSI medium to obtain single colonies. Suspected cured colonies were identified by picking and plating onto TSI medium containing 5 μg/mL of imipenem and onto control TSI medium lacking imipenem. Colonies that failed to grow in the presence of imipenem were suspected to be cured and were further analyzed for blaKPC loss by PCR and antimicrobial susceptibility testing [20].
Whole-genome sequencing (WGS)
We performed WGS of the CPKp1825, CPEa1826, and CPEc1827 isolates using single-molecule real-time sequencing on a PacBio RSII instrument (Pacific Biosciences, Menlo Park, CA, USA). Schematic diagrams of multiple plasmid alignments were prepared by manually realigning the linear plasmid maps drawn using SnapGene Viewer (http://www.snapgene.com/products/snapgeneviewer/). Genomes were compared using Mauve comparison tools (http://darlinglab.org/mauve/). Resistance genes, insertion sequence elements, replication origins, virulent elements, and toxin and antitoxin systems were identified using the following databases: ResFinder (https://cge.cbs.dtu.dk//services/ResFinder/), ISfinder (https://www-is.biotoul.fr/), plasmid finder (https://cge.cbs.dtu.dk/services/PlasmidFinder/), Restriction-Modification finder (https://cge.cbs.dtu.dk/services/Restriction-ModificationFinder//), Serotype finder (https://cge.cbs.dtu.dk/services/SerotypeFinder/), Fim Typer (https://cge.cbs.dtu.dk/services/FimTyper/), and CH Typer (https://cge.cbs.dtu.dk/services/CHTyper/).
GenBank accession numbers
Nucleotide sequence data for CPKp1825, CPEa1826, and CPEc1827 are available under GenBank accession numbers WMHT01000001–WMHT01000003, WMHU01000001–WMHU01000003, and WMHV01000001–WMHV01000003, respectively.
Go to :
RESULTS
Antimicrobial susceptibilities and molecular typing
The Kp (CPKp1825) isolates showed a multidrug resistance (MDR) phenotype. The isolates exhibited resistance to most antimicrobials tested, including ampicillin, aztreonam, cefotaxime, ceftazidime, cefoxitin, ciprofloxacin, gentamicin, tigecycline, imipenem, meropenem, and ertapenem, but were susceptible to amikacin and colistin. K. aerogenes (CPEa1826) and E. coli (CPEc1827) isolates were resistant to ampicillin, aztreonam, cefotaxime, ceftazidime, cefoxitin, imipenem, meropenem, and ertapenem, but were susceptible to amikacin, gentamicin, tigecycline, ciprofloxacin, trimethoprim/sulfamethoxazole, and colistin. The susceptibility test results for combinations of β-lactamases and sites of isolation for the isolates assessed are shown in Table 1.
Table 1
Antimicrobial susceptibilities and epidemiological properties of isolated KPC-producing Enterobacteriaceae*
| Isolate I.D | Specimen | Date | MLST | blaKPC | Plasmid | Strain-susceptible antimicrobials† | Carbapenemase differentiation test | OMP loss | Curing test | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
||||||||||||||||
| ST | Subtype | Bracketed by | pKPC | Replicon | MIC (mg/L) | Zone diameter (mm) | mCIM | eCIM | ||||||||||||
|
|
|
|||||||||||||||||||
| AMK | GEN | CIP | TIG | CST | TMP/SMX | AMK | GEN | CIP | ||||||||||||
| CPKp1825 | Urine | 27-Sep | 307 | blaKPC-2 | ΔTn4401a | pKPBHS25-2 | IncX3 | 16 | 0.25 | 22 | + | - | ompK35, ompK36 | - | ||||||
|
|
||||||||||||||||||||
| CCPKp1825 | blaKPC-2 | ≤ 2 | 0.25 | 22 | ||||||||||||||||
|
|
||||||||||||||||||||
| CPEa1826 | Rectal | 21-Oct | - | blaKPC-2 | ΔTn4401a | pKPBHS26-2 | IncX3 | ≤ 2 | ≤ 1 | ≤ 0.25 | 0.38 | 0.25 | ≤ 20 | 23 | 22 | 30 | + | - | ompK36 | - |
|
|
||||||||||||||||||||
| CCPEa1826 | blaKPC-2 | ≤ 2 | ≤ 1 | ≤ 0.25 | 0.38 | 0.25 | ≤ 20 | 25 | 22 | 30 | ||||||||||
|
|
||||||||||||||||||||
| CPEc1827 | Rectal | 23-Oct | 720 | blaKPC-2 | ΔTn4401a | pKPBHS27-1 | IncX3 | ≤ 2 | ≤ 1 | ≤ 0.25 | 0.125 | 0.25 | ≤ 20 | 20 | 22 | 30 | + | - | ompK35 | - |
|
|
||||||||||||||||||||
| CCPEc1827 | blaKPC-2 | ≤ 2 | ≤ 1 | ≤ 0.25 | 0.75 | 0.25 | ≤ 20 | 23 | 22 | 30 | ||||||||||
* Breakpoints were applied according to the CLSI guidelines [14];
† Disk diffusion test results were interpreted according to the CLSI guidelines [14].
Abbreviations: AMK, amikacin; GEN, gentamicin; CIP, ciprofloxacin; TIG, tigecycline; CST, colistin; TMP/SMX, trimethoprim-sulfamethoxazole; OMP, outer membrane protein; mCIM, modified carbapenem inactivation method; eCIM, EDTA-modified carbapenem inactivation method; CLSI, Clinical and Laboratory Standards Institute.
![]()
PCR and sequencing of β-lactamase genes revealed the presence of blaKPC-2 in all isolates. SHV-1-like broad-spectrum penicillinase, which is intrinsic, was identified by PCR in both CPKp1825 and CPEa1826 isolates. Genes for plasmid-mediated CTX-M-type ESBLs were found in CPKp1825 isolates. In addition, we found that OmpK35 and OmpK36 were missing from CPKp1825 isolates, OmpK36 was missing from CPEa1826 isolates, and OmpK35 was missing from CPEc1827 isolates. Plasmids carrying the blaKPC-2 gene in CPKp1825, CPEa1826, and CPEc1827 could be transferred to E. coli J53. The conjugated strains had the same resistance gene as that of the pre-junction strain (Table 2). A curing test was performed to artificially remove plasmids containing the KPC-2 gene. However, none of the three strains could be successfully cured (Table 1).
Table 2
Resistome and integrative conjugative elements
![]()
Sequencing and annotation of CPKp1825
The Kp strain CPKp1825 has a 5,812,656-bp genome, comprising of a single chromosome and two plasmids (pKPBHS_25_1 and pKPBHS_25_2). The CPKp1825 chromosome consisted of a 5,541,320-bp circular DNA molecule with an average G+C content of 57.2% and harbored 5,143 annotated open reading frames (ORFs). We found no known mutations, but unknown mutations in the genomes included amino acid substitutions P161R, G164A, F172S, R173G, L195V, F197I, and K201M in AcrR. L188Q and K194* substitutions in RamR, OmpK36, and OmpK37 were also found. The type II restriction enzyme M.Kpn34618Dcm was also present. CPKp1825 was identified as an ST307 strain. The chromosome of CPKp1825 isolates included blaSHV-28 and blaSHV-106 for β-lactam resistance, oqxA, B for quinolone resistance, and fosA for fosfomycin resistance.
The multi-replicon (IncFIB (K) and IncFII (K)) plasmid pKPBHS_25_1 included aac (6′)-Ib-cr for fluoroquinolone and aminoglycoside resistance, blaOXA-1 for β-lactam resistance, and catB3 for phenicol resistance (Fig. 1A). Further, pKPBHS_25_1 encoded CusCFBA proteins that mediate resistance to copper and silver by cation efflux, and carried five putative virulence clusters, the lacZYI operon, the Fec-like iron (III) dicitrate and glutathione ABC-transport systems, the urea transport system, and a cluster for glycogen synthesis. A 35-kb region on pKPBHS_25_1 contains a type IV secretion system, the pilus genes, and tra genes for conjugal transfer (Fig. 1B).
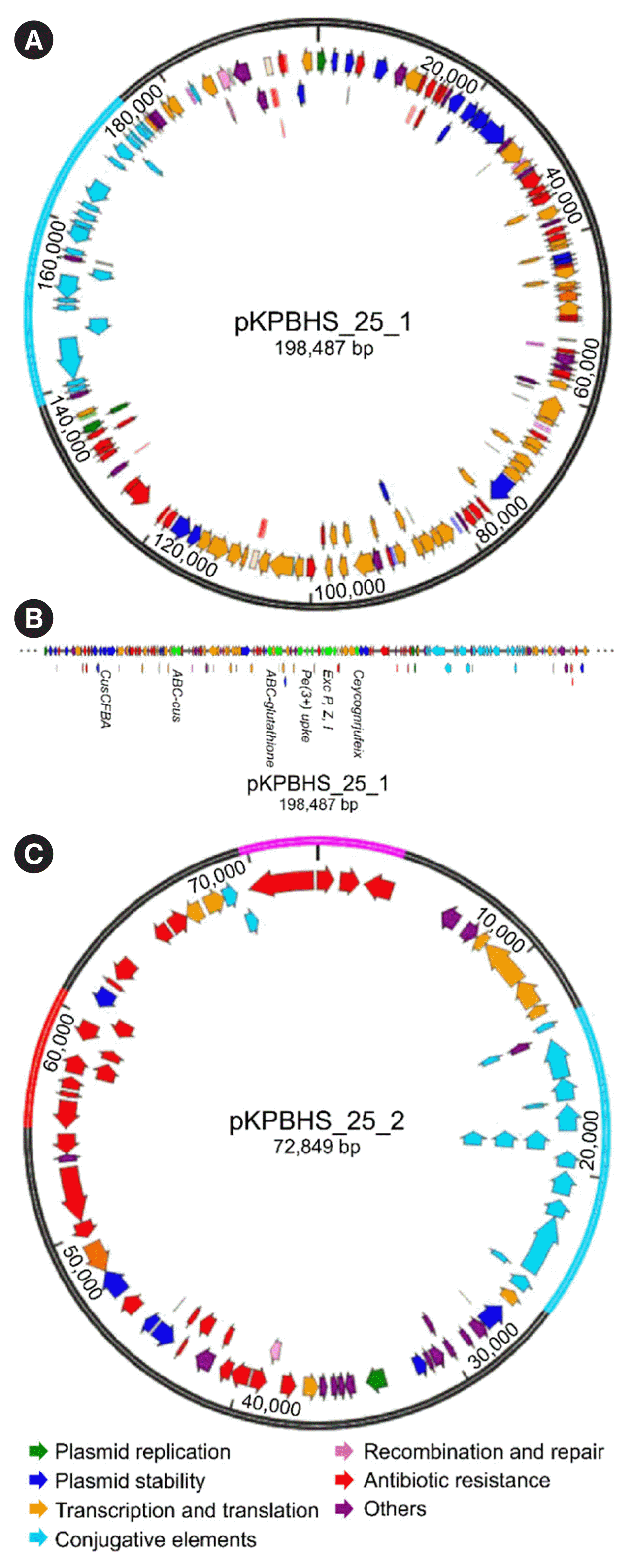 | Fig. 1Genetic organization of plasmids associated with blaKPC. (A) Circular map of pKPBHS_25_1 with two replication origins for the IncFIB (K) and IncFII (K) groups, as well as a copper-transporting efflux system (CusCFBA) and five putative virulence clusters. (B) Variant pKPBHS_25_1 plasmids identified in ST307. (C) Circular map of pECBHS_25_2 containing ΔTn4401a with blaCTX-M-15, blaKPC-2, blaSHV-182, blaTEM-1B, and conjugative elements. Each arrow indicates plasmid scaffold genes and their direction of transcription. The locus Tra is indicated by sky blue arrows, indicating the tra genes (e.g. traG, G; traF, F; traO, O). Related genes (tnpA, tnpR, and tnpM), resistance genes, and insertion sequences are indicated by red arrows. Other genes are indicated by colored arrows as follows: orange, transcription, and translation genes; blue, CusCFBA; green, clusters encoding putative virulence determinants. In the plasmid circle, the Tn3 transposon is indicated in red, ΔTn4401a with blaKPC-2 in pink and conjugative elements in sky blue.
|
pKPBHS_25_2 (72,849 bp), an incompatibility group X type 3 (IncX3) plasmid, was found to belong to an incompatibility group (Fig. 1C) and carry genes encoding type II restriction enzymes Eco128I and M.EcoRII. The plasmid pKPBHS_25_2 contained aac (3)-IIa, aph (3″)-Ib, and aph (6)-Id for aminoglycoside resistance; blaCTX-M-15, blaKPC-2, blaSHV-182, and blaTEM-1B for β-lactam resistance; sul2 for sulfonamide resistance; and dfrA14 for trimethoprim resistance. The blaKPC-2 gene was located on a truncated Tn4401 transposon. ΔISKpn7-blaKPC-2-ISKpn6 had a 99-bp deletion between ISKpn7 and blaKPC, indicating that it is a Tn4401a isoform. The plasmid was found to carry the Tn3 transposon containing blaTEM-1B. A 13-kb region on pKPBHS25-2 contained a type IV secretion system, the pilus genes, and tra genes for conjugal transfer (Fig. 1C and Fig. 2).
 | Fig. 2Linear comparison of the genetic surroundings of blaKPC-2. (A) One fragment unit shows pink, yellow green, and green on the contig. Gene contents were similar along the synteny on the contig. The fragment was well preserved, but inversion existed. Most genes were well preserved. (B) pKPBHS_25_2, pEABHS_26_2, and pECBHS_27_1 are denoted by arrows and colored based on gene-functional classification. Other genes are indicated by colored arrows as follows: green, plasmid replication; blue, plasmid stability; orange, transcription and translation; sky blue, conjugative elements; pink, recombination and repair; red, antimicrobial resistance; purple, other genes.
|
Sequencing and annotation of CPEa1826
WGS revealed that the K. aerogenes strain CPEa1826 has a 5,254,026-bp genome with two plasmids (pEABHS_26_1 and pEABHS_26_2). The nucleotide sequence variations of unknown mutations in 23S, 16S rsH, B, C, and amino acid change of rpoB were confirmed. M.SenAnal, the methyltransferase type I restriction enzyme, was also detected. The chromosome CPEa1826 did not contain any acquired antimicrobial resistance determinants. Plasmid pEABHS_26_1, belonging to the IncP-1 group, did not contain any acquired antimicrobial resistance gene, and encoded duplicated type IV secretion systems and conjugative elements. The 46,836-bp plasmid pEABHS_26_2 carrying the blaKPC-2 and blaSHV-182 genes belongs to the IncX3 group. pEABHS_ 26_2 had an average G+C content of 48.0% and harbored 65 annotated ORFs. The blaKPC-2 gene was located on a truncated Tn4401 transposon (Fig. 2 and Fig. 3A).
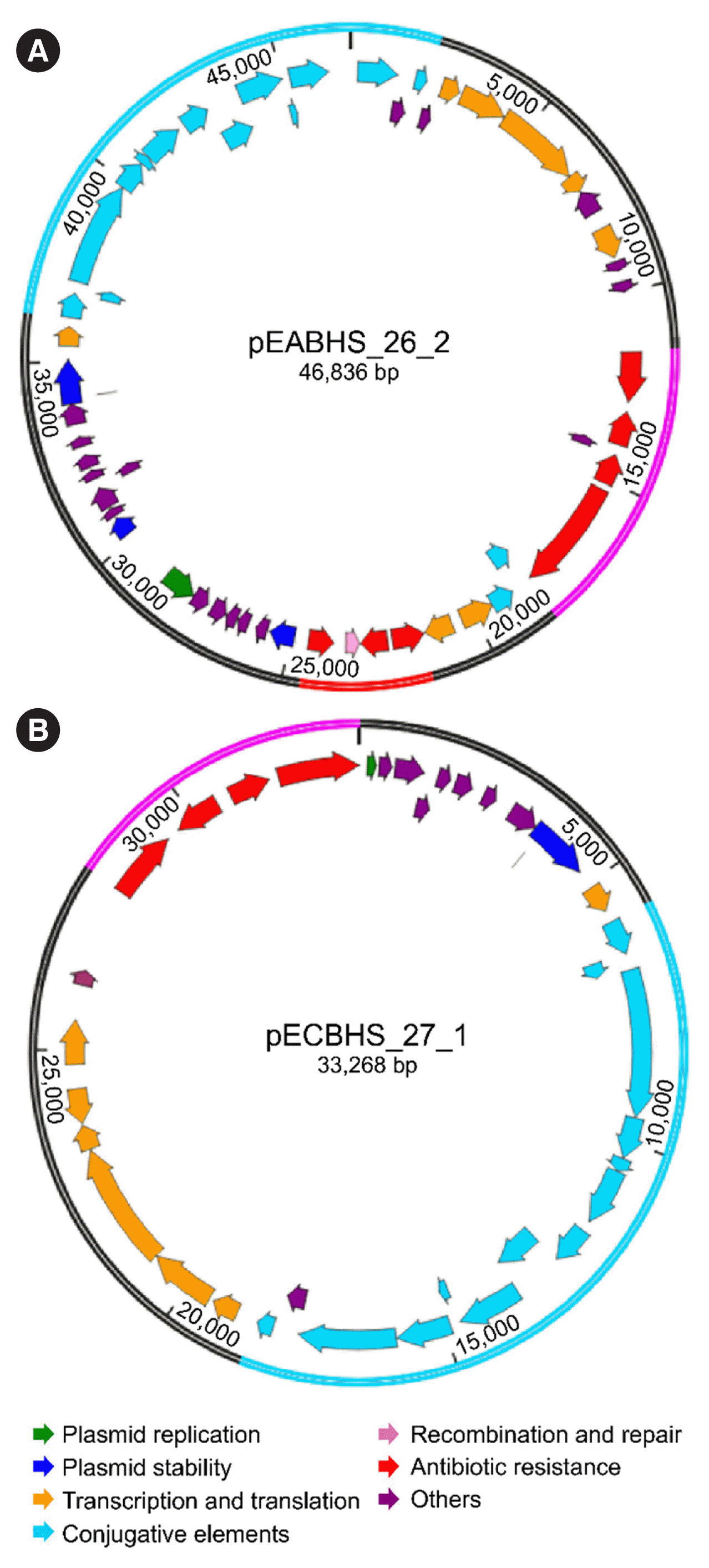 | Fig. 3Genetic organization of plasmids pEABHS_26_2 and pECBHS_27_1. (A) Circular map of pEABHS_26_2 containing ΔTn4401a harboring blaKPC-2 and blaSHV-182 genes. (B) Circular map of pECBHS_27_1 containing ΔTn4401a harboring blaKPC-2 and conjugal transfer genes. In the plasmid circle, the Tn3 transposon is indicated in red, ΔTn4401a with blaKPC-2 in pink and conjugative elements in sky blue.
|
Sequencing and annotation of CPEc1827
The E. coli strain CPEc1827 was found to have a 5,033,400-bp genome comprising a single chromosome and a plasmid. The genome had a S57T substitution in ParC. The nucleotide sequence variations of unknown mutations of 16S_rrsB, H, C, and 23S, and amino acid changes in pmrB and parC, as well as the nucleotide sequence variations of unknown mutations of the 23S, 16S rsH, B, C, and amino acid changes in rpoB were confirmed. Type II restriction enzymes M.EcoE455Dcm and M.EcoGVI were also found. CPEc1827 was identified as ST720 and belonged to the serotypes ST17/O77 and H41, while its subtype to fum C3 and fim H65. The chromosome included mdf (A) for macrolide, lincosamide, and streptogramin B resistance. The 91,356-bp contig did not contain any acquired antimicrobial resistance determinants. The 33,268-bp IncX3 plasmid (pECBHS_27_1) carried blaKPC-2 for β-lactam resistance (Fig. 3B). The blaKPC-2 gene was found to be located on a truncated Tn4401 transposon, and conjugal transfer genes (including tra and trb) were located at 12,738 bp (Fig. 2 and 3B).
Go to :
DISCUSSION
KPC-Kp ST307 is a novel lineage that has the potential to become an epidemic or ‘high-risk’ clone. ST258 is a well-known KPC-producing clone with a global distribution [3, 4]. However, according to the literature, it is not prevalent in Korea [21]. The most common clone in Korea is KPC-Kp ST307. Several reports have indicated local dissemination of ST307 harboring the blaKPC genes blaKPC-2 (Columbia, USA, Korea) and blaKPC-3 (Italy) [8, 22, 23], whereas an analysis of 1,700 ESBL-producing Kp from a hospital network in Texas, USA revealed a high prevalence of blaCTX-M-15-positive ST307 strains, with one-third of the strains also carrying blaKPC-2 genes and only three carrying blaKPC-3 [24].
The blaCTX-M-15 gene is common in ST307 strains [21, 24, 25]. According to Wyres, et al. [26], ParC 80I and GyrA 83I fluoroquinolone resistance-associated mutations were conserved in all 95 genomes analyzed in their study. The study also showed the blaCTX-M-15 ESBL gene to be present in 89 genomes (93.7%), and 81 genomes (85.3%) harbored sul2, dfrA14, and strAB with or without aac(3)-IIa, which were all linked to an MDR plasmid. These results are consistent with our findings. In addition to blaCTX-M-15, blaSHV-182 and blaTEM-1B were repeatedly detected during the 38-day hospitalization period of the patient, resulting in continuous infection. Thus, plasmid-mediated glycogen synthesis might help ST307 isolates survive under limited nutrient availability, and the urea transport system might facilitate colonization of this strain in the urinary tract.
Type II restriction enzymes were observed in CR-KP1725 and CR-KP1727. Type II restriction enzymes function to protect their host DNA by removing extracellular foreign DNA that enters the cell to maintain its pathological state. The presence of different promoters, leading to various levels of expression of the type II restriction enzymes, could be responsible for the variability in resistance levels between CR-KP1725 and CR-KP1727, but this might not be sufficient to explain the high level of resistance, as previously suggested [27].
All three strains had lost their outer membrane proteins. The strain lacking OmpK36 alone became resistant to cefazolin, cephalothin, and cefoxitin, indicating that the disruption of ompK36 results in greater drug resistance than the disruption of ompK35. The additional loss of OmpK35 further increased the MICs, which demonstrated that the double-deletion strain was highly resistant to the antimicrobials [28]. All these genetic functions synergistically make KP ST307 more resistant to antimicrobials and more difficult to treat in patients. The MIC results for amikacin in this study should be interpreted cautiously [29]. Sensitivity was confirmed in MIC and antimicrobial disk tests, but WGS analysis revealed that genes including aac (3)-lla, aph (6)-ld, and aph (3″)-lb were resistance genes. Only 3% (2/65) and 11% (7/65) of the isolates were non-susceptible to amikacin using the CLSI and EUCAST breakpoints, respectively [14, 30]. These low rates were observed even though 73% (16/22) of aminoglycoside-modifying enzyme (AME)+isolates possessed aac (6′)-Ib, which is reported to confer amikacin resistance. Amikacin MICs below the susceptibility breakpoints are commonly observed among Enterobacter and other Enterobacteriaceae that possess aac (6′)-Ib [29, 31]. The clinical significance of low MICs against bacteria carrying AMEs that are known to cause resistance has not been established, and it is unclear whether MICs or molecular markers are more relevant to treatment responses [32]. Indeed, the EUCAST states that should a member of the Enterobacteriaceae test as tobramycin (TOB)-intermediate or TOB-resistant and gentamicin- and amikacin-susceptible, its amikacin susceptibility status should be revised to “intermediate” [30]. Until the clinical significance of these low MICs is resolved, we recommend that amikacin be used with caution in the treatment of infections caused by Enterobacter that are resistant to other aminoglycosides.
Our study revealed that the blaKPC-2 gene from KPC-Kp ST307 could be horizontally transferred to other bacterial species. The blaKPC-2 gene of KPC-Kp ST307 is encoded by a conjugable plasmid, which can be speculated to provide a possible mechanism for dissemination to other species of gram-negative bacilli. Plasmids from all three strains could be transmitted through conjugation. Tn4401, a transposable element, could also move across the species. This shows the limitation of symptomatic treatment, as well as the risk posed by KPC-Kp ST307. CR Enterobacteriaceae-infected patients have a high mortality rate, and the therapeutic agents used to treat their condition are limited. Thus, treatment of CR Enterobacteriaceae infection often involves a combination therapy, including carbapenem, aminoglycosides, and fosfomycin. Monotherapies of colistin or tigecycline are the last line of treatment; however, this therapeutic option is not yet available in Korea.
This study had two limitations. First, only KPC-producing Enterobacteriaceae strains observed at admission were analyzed, and molecular microbiological evidence was not generated for carbapenemase resistance in Enterobacteriaceae infections that occurred later, which limits the generalizability of our findings. Second, an epidemiological study of the KPC-Kp isolates to identify their origin of occurrence was not conducted. However, the ease of horizontal transfer of KPC-Kp ST307 plasmids observed in this study has serious public health and epidemiological implications. MDR Kp ST307 should be monitored alongside other well-known clones to detect emerging MDR threats. Our findings indicate an urgent need for enhanced surveillance of KPC-Kp ST307. In particular, the occurrence of KPC-Kp ST307 in Korea, with its high frequency, suggests a health threat. Controlling the spread and limiting the impact of KPC-Kp ST307 isolates in Korean hospitals will require intensive efforts in both public and private healthcare centers. These data may serve an important role in understanding and controlling the spread of CR gram-negative pathogens.
Go to :
Supplementary Information
Supplemental Data Table S1
Primer for PCR identification of resistance determinants.
Go to :
 XML Download
XML Download