

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Spinal muscular atrophy (SMA) is the most common autosomal recessive neurodegenerative disorder resulting from loss of motor neurons in the spinal cord and brainstem, leading to hypotonia, progressive proximal muscle weakness, and respiratory distress. SMA has a reported incidence of approximately 1 in 7,000 live births [1] and a high carrier rate ranging from 1.38% to 2.13% [2, 3]. This severe neuromuscular disease has been recommended for routine carrier screening in the general population by the American College of Medical Genetics [4, 5].
Approximately 95% of SMA patients show a homozygous absence of exon 7 or both exons 7 and 8 of the survival of motor neuron 1 (SMN1) gene, which is located on chromosome 5q13 [6]. This region contains two nearly identical copies: the telomeric copy (the SMN1 gene) and the centromeric copy (the SMN2 gene), which increases the difficulty of molecular diagnosis of SMA by detecting SMN1 exon 7 copy number. The SMN1 gene differs from the SMN2 gene by only five single nucleotides including four in the non-coding region and one in the coding region (exon 7) [7]. The only coding region variant, a single C-to-T substitution between SMN1 and SMN2 in exon 7 (c.840C>T), results in a decrease in full-length SMN2 transcripts and is considered to mainly differentiate SMN1 from SMN2.
Real-time PCR and PCR-denaturing high performance liquid chromatography (PCR-DHPLC) are the most common technologies for SMA large-scale population screening [8–10]. However, the primers or probes used in traditional SMA PCR strategies, which cover the single differential base pair, may not fully ensure SMN1 amplification specificity. Recent studies have shown that oligonucleotide sequences modified by a locked nucleic acid (LNA), an available alternative nucleic acid that contains a methylene bridge connecting the 2′oxygen and 4′-carbon in the ribose moiety, display enhanced specificity, sensitivity, and hybridization stability toward complementary DNA and RNA [11, 12]. Substitution with an LNA monomer increases primer melting temperature (Tm) values from +2 to +8°C for DNA, which may be related to the increased thermal stability of the base pairs. Additionally, the increase in Tm value depends on the position and number of the LNAs in oligonucleotide chain [11].
Therefore, to further optimize a real-time PCR approach for SMN1 detection, we modified conventional primers and probes with LNAs and validated the test performance using clinical samples. We also expanded the application of this new technology for SMA screening in newborns using dried blood spots (DBS). To the best of our knowledge, this is the first duplexed real-time PCR approach developed using LNA-modified primers for detecting the copy number of SMN1 exon 7.
MATERIALS AND METHODS
Sample sources
In this prospective study, 157 peripheral blood samples (40 patients, 74 carriers, and 43 normal individuals), whose genotypes were confirmed by multiplex ligation-dependent probe amplification (MLPA) at Hunan Jiahui Genetics Hospital, Hunan, China, from May 2014 to December 2018, were collected. Of these, 60 samples were used to establish the real-time PCR test, while 97 samples (20 patients, 54 carriers, and 23 normal individuals) and another 25 DBS (derived from peripheral blood) with genotypes verified using MLPA were randomly assigned in a double-blind design. Genomic DNA was extracted from peripheral blood cells using the QuickGene DNA whole blood kit L and the QuickGene-610L automated system (FUJIFILM Corporation, Tokyo, Japan) and kept at −20°C. Since 2018, 753 newborn DBS samples from the Han population preserved at −20°C have been randomly selected from the Maternal and Child Health Hospital of Hunan Province, China, for SMA screening. Genomic DNA was extracted from the DBS using the Nucleic Acid Extraction Reagent (Shenzhen Uni-medica Co., Ltd., Shenzhen, China). Our study complies with the Declaration of Helsinki and was approved by the Institutional Review Board of the Center for Medical Genetics, Central South University, Hunan (201503002). Informed consent was obtained from individuals and parents of the newborns involved in the study.
Primers and probes
The primers and probes were designed using Primer Premier 5 (Premier Biosoft, San Francisco, CA, USA). The 3′ ends of the specific forward primer (SMN1-F) and reverse primer (SMN1-R) used for amplifying the SMN1 gene (NM_000344.3) were designed to exactly target the c.835-44g and c.840C sites, respectively, and were modified by LNAs (Fig. 1). To further prevent nonspecific amplification of the SMN2 gene (NM_017411.3), we also designed a blocked probe (SMN2-B) targeting the c.840T site; the 18th and 19th bases of this probe were modified with an LNA, and the 3′ end was modified with a C3 spacer. The primers for the control gene, albumin (ALB; NM_000477.7), were designed in the region between intron 11 and exon 12. Reporter dyes 6-carboxyfluorescein (FAM) and cyanine 5 (Cy5) were used to label the TaqMan probes of SMN1 and ALB, respectively. To further optimize the amplification specificity for SMN1, we designed two primer pairs: nonspecific primers SMN-F/R and specific primers SMN1-F1/R1 without LNA modification. All primers and probes (Supplemental Data Table S1) were synthesized by Sangon Biotech (Shanghai, China). Sanger Sequencing with different primer combinations was also conducted by Sangon Biotech (Fig. 1).
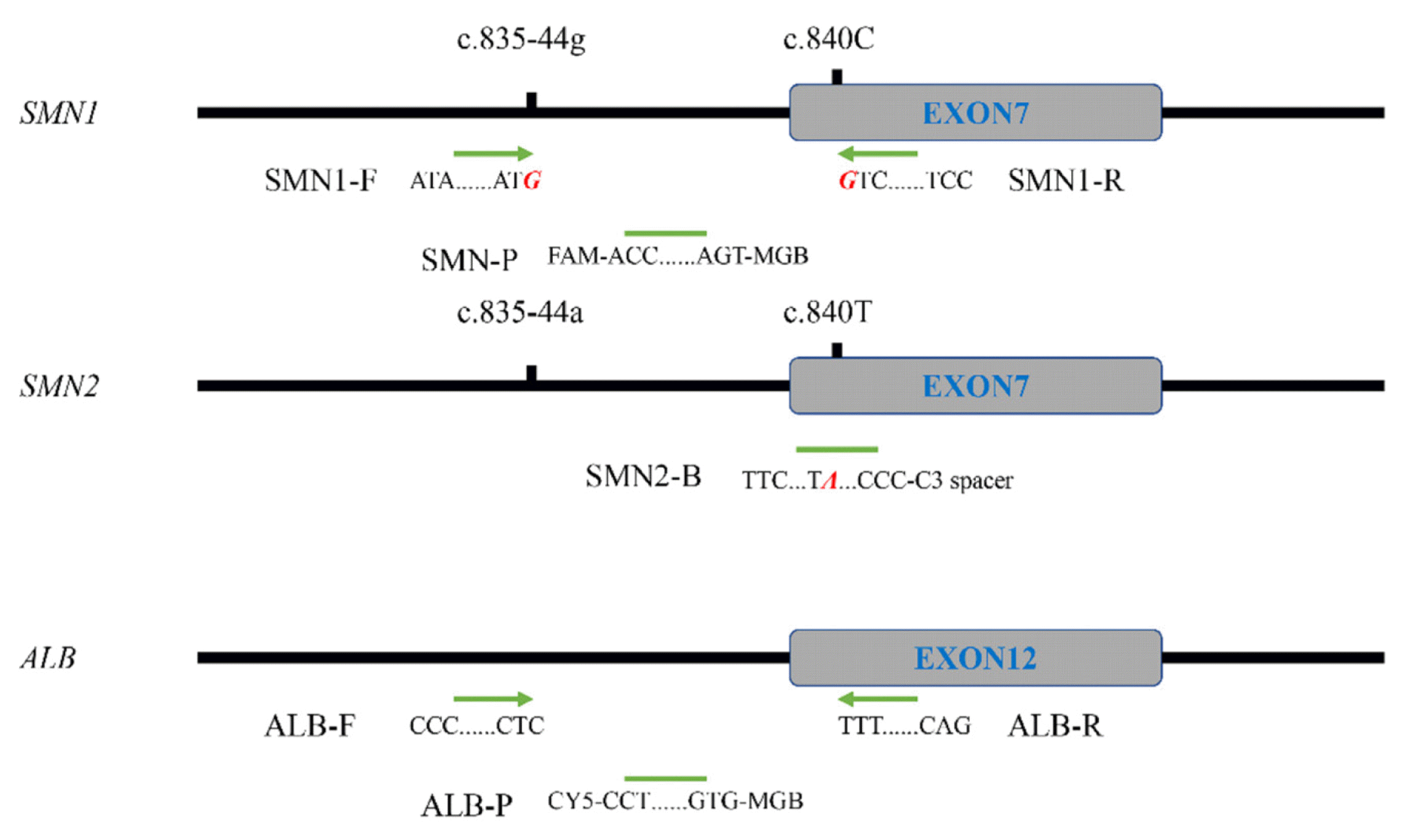
Fig. 1
Diagram of the binding location of the SMN1, SMN2, and ALB primers and probes. In primers SMN1-F/R, the 3′ end of the forward primer and reverse primer, targeting the c.835-44g and c.840C site, respectively, has been LNA-modified; the modified bases are indicated in red.
Abbreviations: SMN, survival motor neuron; ALB, albumin; LNA, locked nucleic acid; F, forward; R, reverse; P, probe.
![]()
MLPA
A total of 157 peripheral blood samples were analyzed using MLPA to confirm the copy number of exon 7 and 8 in the SMN1 and SMN2 genes using the SALSA MLPA P021 SMA kit (MRC-Holland, Amsterdam, The Netherlands).
Real-time PCR
Real-time PCR was performed using an ASA-9600 Real-Time PCR System (Suzhou Baiyuan Gene Technology Co., Ltd., Suzhou, China) with a 2× AceQ U+ Probe Master MIX (Vazyme Biotech Co., Ltd., Nanjing, China) in triplicate and run according to the manufacturer’s protocol. We initially used a temperature gradient (from 54–60°C) to determine the optimal PCR annealing temperature for maximal and stable amplification efficiency. PCR was then performed with the following amplification conditions: 45°C for 2 minutes and 95°C for 10 minutes, followed by 45 cycles of 95°C for 15 seconds and 60°C for 40 seconds. Partial DNA sequences of SMN1 and ALB were cloned into a single pUC57 vector as the standard samples for quantifying SMN1 and ALB copy number (Supplemental Data Fig. S1). To facilitate test interpretation, we defined three rate value (R) ranges according to the relative rate between the copy number of SMN1 and ALB after one duplexed reaction. Based on the previous experiment with 60 samples, patients, carriers, and normal individuals were assigned ratios of 0–0.1, 0.35–0.65, and 0.8–1.2, respectively. To validate clinical efficacy, another 97 clinical DNA samples extracted from peripheral blood were recruited and tested by randomized double-blind design using the established duplexed real-time PCR approach.
RESULTS
Primer specificity was increased by LNA modification
As the homozygous deletion of SMN1 exon 7 in the patient DNA was previously confirmed, increased PCR products would indicate increased nonspecific amplification of pseudogene SMN2. Accordingly, the real-time PCR results showed that the internal control ALB amplification and targeted gene amplification with complete nonspecific primers SMN-F/R entered the exponential phase at the same time, indicating complete nonspecific SMN2 amplification. When the forward or reverse primer was replaced with the allele-specific primer SMN1-F1 (c.835-44g) or SMN1-R1 (c.840C), respectively, the exponential phase was obviously delayed, suggesting less SMN2 amplification. Thus, although both allele-specific primers exhibited high specificity for SMN1, they could not eliminate the nonspecific amplification of SMN2. When the allele-specific primers were further modified with LNAs, no SMN2 amplification was detected with either a single LNA-modified primer or both LNA-modified primers (SMN1-F/R); the amplification curves did not form or remained under the threshold line in valid cycles, implying an extreme enhancement of SMN1 specificity following LNA modification (Fig. 2). A similar increase in specificity due to LNA modification was observed in the Sanger sequencing chromatograms of the two SMN1 and SMN2 differential sites (Supplemental Data Fig. S2).
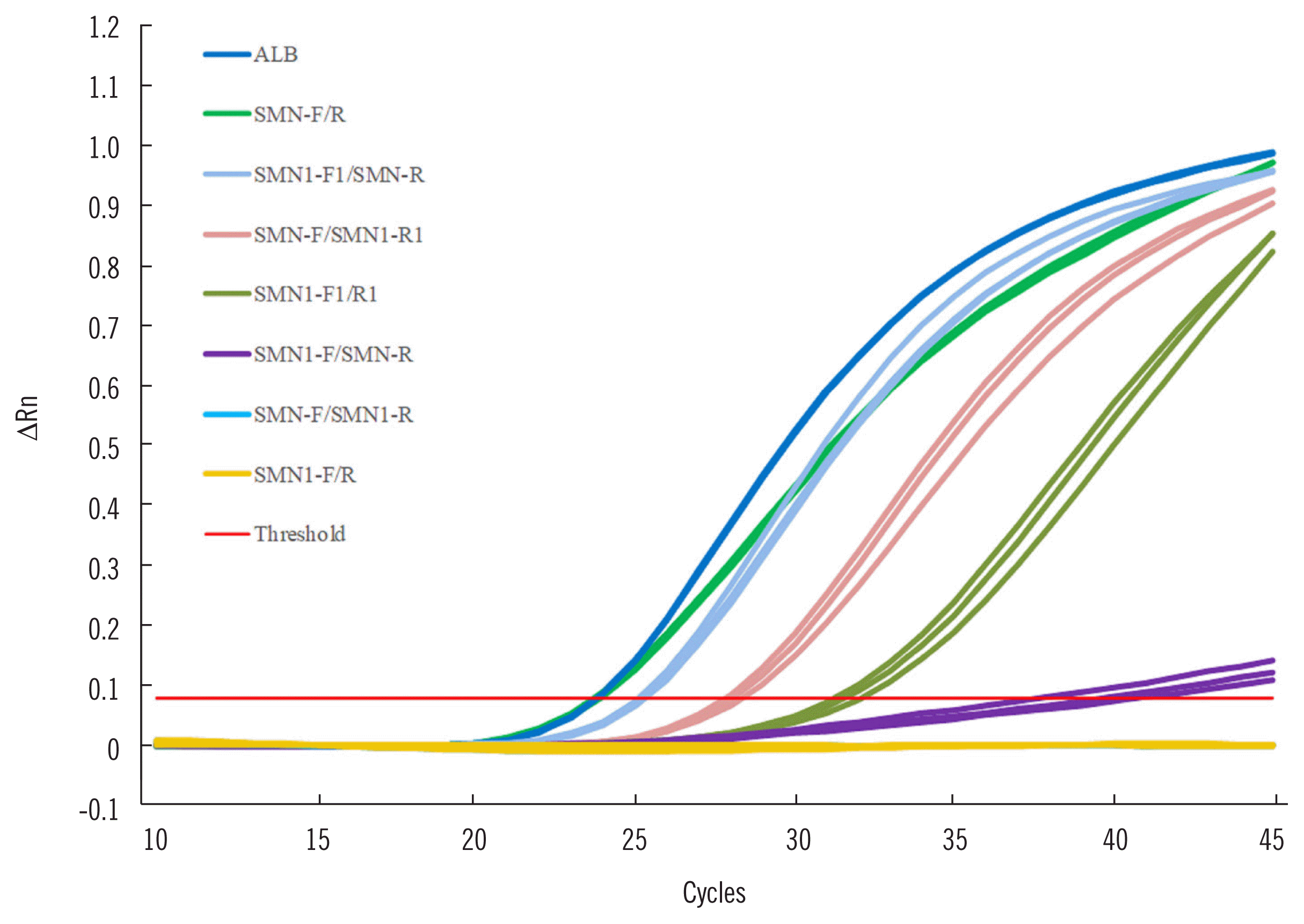
Fig. 2
The amplification specificity of seven different SMN1 primer groups detected by real-time PCR using SMA patient DNA as the template. Each group was set up in triplicate.
Abbreviations: SMN1, survival motor neuron 1; ALB, albumin; SMA, spinal muscular atrophy; ΔRn, normalized reporter; F, forward; R, reverse.
![]()
Validation using clinical peripheral blood and DBS samples
The results revealed a uniform efficiency of approximately 95% throughout the temperature range (54–60°C) when using normal individual DNA samples with the SMN1-F/R primers, which was 6.8% higher than when primers SMN1-F1/R1 (without LNA modification) were used at 60°C (Fig. 3A & 3B). Hence, LNA modification may not only improve primer specificity but also promote amplification efficiency. The three genotypes are clearly identifiable on the basis of the amplification curve. Normal individual samples showed a synchronous cycle threshold (Ct) with ALB, indicating two copies of SMN1 exon 7 (Fig. 3C), while the carrier samples displayed a 1 Ct extension compared with ALB, suggesting one copy of SMN1 exon 7 (Fig. 3D). No amplification of SMN1 was observed in SMA patient samples, confirming homozygous deletion of SMN1 exon 7 and no detectable cross-reactivity with SMN2 (Fig. 3E). These 60 test results demonstrated 100% concordance with the corresponding clinical status. ALB demonstrated stable amplification in all samples in the duplexed test with nearly the same Ct (Fig. 3).
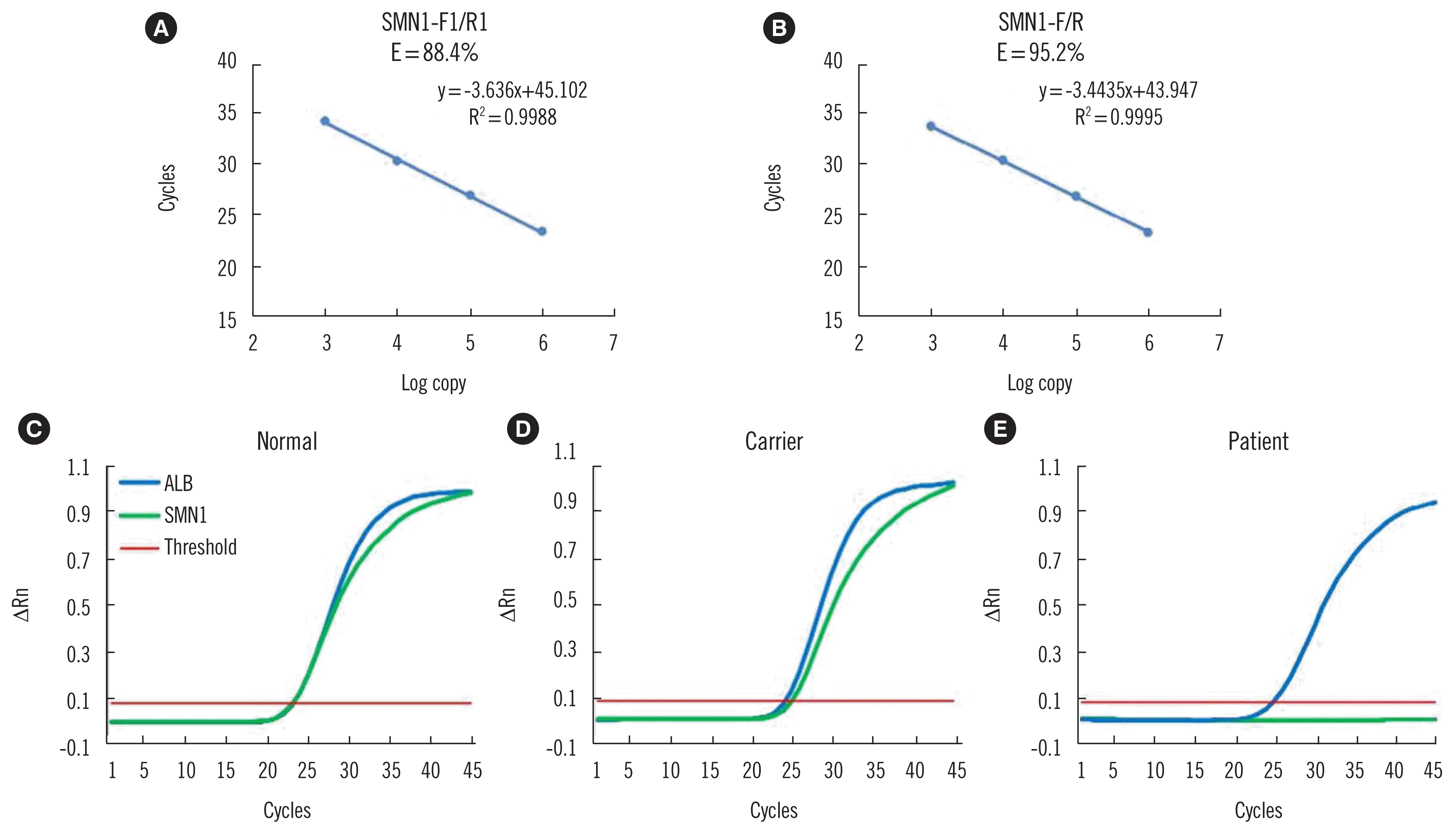
Fig. 3
Primer amplification efficiency and real-time PCR amplification curves. (A) The amplification efficiency of primers SMN1-F1/R1 was 88.4% at 60°C. (B) The amplification efficiency of primers SMN1-F/R was 95.2% at 60°C. (C) The SMN1 and ALB amplification curves in normal individual samples overlapped at the threshold. (D) In SMA carrier samples, the SMN1 amplification curve exhibited an approximate one cycle delay compared with the ALB curve at the threshold. (E) In SMA patient samples, only the ALB gene was amplified.
Abbreviations: SMN1, survival motor neuron 1; ALB, albumin; SMA, spinal muscular atrophy; F, forward; R, reverse; ΔRn, normalized reporter.
![]()
Of the 97 samples, 20 were considered to be SMA patients (average R: 0, R range: 0–0), 54 were classified as SMA carriers (average R: 0.56±0.03, R range: 0.49–0.62), and the remaining 23 samples were classified as normal individuals (average R: 1±0.09, R range: 0.8–1.18). After unblinding the trial, all the test results were completely consistent with their original phenotypes and genotypes, which were confirmed by MLPA. Therefore, the sensitivity and specificity of our new real-time PCR technology was 100% for SMA patients with no copies of SMN1, carriers with one copy of SMN1, and normal individuals with two copies of SMN1 (Table 1).
Table 1
Validation using 97 peripheral blood samples by real-time PCR
| SMN1 copy number | |||||
|---|---|---|---|---|---|
|
|
|||||
| 0 | 1 | 2 | 3 | Total | |
| Individuals | 20 | 54 | 23 | 0 | 97 |
| R-value* | 0 | 0.56±0.03 | 1±0.09 | - | - |
|
|
|||||
| Sensitivity | 100% | 100% | 100% | ||
| 95% CI | 80–100% | 92–100% | 82–100% | - | - |
| Specificity | 100% | 100% | 100% | ||
| 95% CI | 94–100% | 90–100% | 94–100% | - | - |
![]()
To expand the application of this new approach to newborn screening for SMA, we also performed a double-blind test using 25 DBS samples (5 SMA patients, 10 SMA carriers, and 10 normal individuals). The SMN1 categorical genotypes obtained from the test also showed 100% concordance with the clinical status of the samples (Table 2).
Table 2
Detection of 25 DBS derived from peripheral blood and screening of 753 newborn DBS samples using real-time PCR
![]()
Newborn screening for SMA patients and carriers using the duplexed approach with DBS
Of 753 newborn DBS samples, 15 newborns were identified as SMA carriers with one copy of SMN1 (average R: 0.54±0.06, R range: 0.43–0.63) and 729 were identified as normal individuals with two copies of SMN1 (average R: 0.97±0.12, R range: 0.8–1.2). Surprisingly, nine newborns were identified as having three copies of SMN1, with an R-value of approximately 1.5 (average R: 1.49±0.08, R range: 1.38–1.61). These data demonstrate, for the first time, a carrier rate of 1.99% (95% confidence interval, 1.12–3.26%) in Hunan Province, South Central China (Table 2).
DISCUSSION
The high carrier rate and severe clinical outcome of infantile SMA necessitates performing timely carrier screening for pregnant women and couples with a birth plan [13, 14]. In addition, as two US Food and Drug Administration approved drugs for SMA treatment have entered clinical practice [15], newborn screening for SMA by genetic testing would be an effective measure for maximizing the benefits of presymptomatic intervention and guide subsequent healthcare.
Although several technologies, such as MLPA, droplet digital PCR, and next generation sequencing (NGS)-based approach, display high specificity and sensitivity for SMA detection [16,17], relatively complex processes, equipment requirements, and high costs currently obstruct their application in population screening, especially for grass-root units. PCR-based approaches remain preferable options for large-scale population screening. The main challenge of conventional PCR-related approach in detecting SMN1 absence is the potential nonspecific amplification of SMN2, owing to the nearly identical sequence of SMN1 and SMN2.
In accordance with the allele-specific PCR principle, when we performed the real-time PCR approach using SMA patients’ DNA with conventional PCR primers, nonspecific amplicons of SMN2 were detected, indicating that conventional real-time PCR may have a potential risk of producing false-negative signals. To reduce this risk, we introduced LNA modification in the primers to further promote the specificity of SMN1 amplification; LNA oligonucleotides increase primer or probe binding strength for complementary sequences, thereby enhancing test specificity and enabling detection of single nucleotide mismatches [18, 19]. A previous SMA detection study reported that a five LNA monomer-modified probe, targeting the differential site c.888+100a in the SMN1 noncoding region, markedly increased detection specificity. When the annealing temperature raised to 65°C, no nonspecific amplicons were detected using the LNA probe in SMA patients [20]. However, that technology was not sufficiently precise for reliable carrier identification. Additionally, another study using a similar probe targeting c.888+100a, reported eight false-positive cases [21], which may also cause false-negative results owing to the conversion of SMN1 into SMN2 following a c.840C>T substitution. We utilized two differential sites in SMN1, c.835-44g and c.840C, at the 3′ end of our designed primers. Additionally, in contrast to the five LNA monomer-modified probe, we used only one LNA to modify the 3′ end of the designed primers, consequently obtaining a significant increase in specificity.
Furthermore, LNA-modified primers appear to enhance the stability and sensitivity of SMN1 detection in a duplexed test. Our results revealed that although the duplexed test contained complex components, including four primers and three probes, as well as different templates, stable and satisfactory amplification efficiency was maintained even when the annealing temperature was changed from 54 to 60°C. Subsequent clinical efficacy demonstrated 100% concordance with each group of patients, carriers, and normal individuals.
A cohort of 753 newborn DBS samples screened using our novel duplexed real-time PCR approach, demonstrated, for the first time, a carrier rate of 1.99% among the Han population of Hunan Province. This result is similar to the carrier rates of 1.90% and 1.83% revealed by studies conducted in East China [22] and Korea [23], respectively.
The cost for our new approach is approximately $1 dollar per test, and the results are available within three hours. Although approximately 95% of SMA patients with a homozygous absence of exon 7 and corresponding carriers can be detected using this economical and rapid test, a small number of patients with SMA caused by single nucleotide variation and (2+0) carriers (with two copies of SMN1 on the same chromosome) cannot be identified [24, 25]. Further efforts should focus on the development of more comprehensive screening approaches for SMA.
In conclusion, we have developed a duplexed real-time PCR approach using LNA-modified primers for detecting the copy number of SMN1 exon 7. The approach is suitable for newborn screening with trace DNA samples, such as DBS, and for carrier screening of pregnant women using peripheral blood DNA.
Supplementary Information
Supplemental Data Fig. S1
The SMN1 (blue) and ALB (red) sequences that were cloned into a single pUC57 vector as the standard samples.
Abbreviations: SMN1, survival motor neuron 1; ALB, albumin.
Supplemental Data Fig. S2
The specificity of SMN1-F and SMN1-R was confirmed by Sanger sequencing. (A) The height of the c.840C/T sequencing peak was used to verify the specificity of primers SMN1-F1 and SMN1-F. The PCR products of non-specific primers SMN-F and SMN-R exhibited nearly equal height C/T peaks. A low T peak appeared in the PCR product of non-specific primer SMN-R and specific primer SMN1-F1 (without LNA modification). A single C peak appeared in the PCR product of non-specific primer SMN-R and specific primer SMN1-F (with LNA modification), indicating complete elimination of nonspecific amplification. (B) The height of the c.835-44g/a sequencing peak was used to verify the specificity of primers SMN1-R1 and SMN1-R. The PCR products of non-specific primers SMN-F and SMN-R exhibited nearly equal height g/a peaks. A single g peak appeared in the PCR products of non-specific primer SMN-F and specific primer SMN1-R1 or SMN1-R (without or with LNA modification), indicating that both specific reverse primers exhibit high specificity for SMN1.
Abbreviations: SMN1, survival motor neuron 1; LNA, locked nucleic acid; F, forward; R, reverse.
 XML Download
XML Download