

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Successful outcome in adhesive dentistry means long-lasting restorations. However, there is considerable evidence that this target is not always reached and a large amount of research work has been carried out in aiming to improve the durability of resin-dentin bonding [1].
The integrity of the adhesive interface does not remain constant mainly due to the hydrolytic degradation of the adhesive monomers in a hybrid layer over a period of time and inability of an adhesive to fully permeate and cover the collagen mesh. Those unprotected demineralized collagen fibrils underlying the hybrid layer are exposed to proteolysis by host enzymes [2,3].
Dentin is a mineralized tissue composed of inorganic apatite crystals embedded in an extracellular organic matrix (EOM). Type I collagen is the main component of the EOM, accounting for up to 90% of the organic material. In addition, various proteins collectively referred to as non-collagenous proteins constitute approximately 10% of the matrix and include proteoglycans, phospholipids and enzymes. Among dentin enzymes, matrix metalloproteinases (MMPs) have recently gained much attention because of their possible role in various physiological and pathological processes in dentin [4]. MMPs are zinc- and calcium-dependent endopeptidases involved in tooth development, dentin caries progression and hybrid layer degradation in composite resin restorations [5].
Bonding of resinous materials to dentin requires prior application of acid conditioners to promote a demineralized surface and expose the organic dentin matrix. In addition, the surface has to be fully infiltrated by monomers from the adhesive system, which will be polymerized to form a tooth–restoration interface called the hybrid layer [6]. However, this mechanism is not fully effective and collagen fibrils remain unprotected in some areas where monomer infiltration is deficient and are susceptible to the activity of MMPs [7].
Under physiological conditions, these proteases are secreted by odontoblasts during dentinogenesis and remain entrapped within the extracellular matrix of dentin in inactive form after matrix mineralization. The acid environment resulting from caries processes, including demineralization with phosphoric acid or acid resinous monomers derived from self-etching adhesives, activates different dentin MMPs [8,9] that are responsible for the degradation of components of the extracellular matrix, mainly collagen type I [9,10].
MMPs play an important role in the degradation process of the hybrid layer, leading to loss of strength in the resin-dentin bond, a problem that directly influences the longevity of adhesive restorations [7]. Thus, the inhibition of MMPs may represent a key piece of the troubleshooting for dentin adhesion, and this literature review aims to bring an overview of the strategies to inhibit MMPs and achieve a long-lasting of the hybrid layer.
Literature search was performed comprehensively using the electronic databases: PubMed, ScienceDirect and Scopus from May 2007 to October 2018. The main search terms were “matrix metalloproteinases”, “collagen”, and “dentin” and “hybrid layer”. These keywords and terms were selected from key articles published in Medical Subject Headings (MeSH). They were enriched with additional terms during the electronic database searches. Articles published in English language were included. The search strategy was modified for each electronic database. A hand search of the reference lists of eligible articles was also performed to identify relevant articles. The citations thought to fulfill the inclusion criteria described above were assessed as full articles. Only articles published in the peer-reviewed journals were included. Although the review comprised primarily in vitro studies, in vivo studies were also included. From the selected articles evaluation of their cited references yielded additional papers suitable for inclusion in this review.
Go to : 
REVIEW
Hydrolytic degradation of adhesive polymers
Adhesive systems are applied on a water-filled collagen network created after acid etching the dentin to infiltrate the spaces to form a hybrid substrate that is the bonding interface between tooth/restorative materials. To reach this aim, dentin should be kept hydrated, preventing the collapse of collagen fibrils. Water within the hybrid layer serves as a functional medium for the hydrolysis of the resin matrix. Hydrolysis and leaching of resin adhesives are considered the primary reason for resin degradation within the hybrid layer, contributing to the reduction of bond strength overtime [1]. Despite their hydrophobic characteristics, the resinous adhesives absorb water over time. This water plasticizes the polymer chains by acting as a molecular lubricant. The hydrophilic portion of the adhesives and the fluids sorption by material allows the flow of water, which forms large aqueous channels within the hybrid layer that work as the pathway to leachable unpolymerized monomers. The presence of water promotes the chemical hydrolysis of ester bonds in methacrylate materials. After that, local domains of the methacrylate network may get sufficiently degraded and/or hydrophilic to permit access of esterases, which greatly accelerate ester-bond hydrolysis [1,11]. This phenomenon can be triggered not only with etch-and-rinse adhesives but also with self-etching adhesives [6,12].
Collagen type I: structure and importance for bond stability
The type I collagen molecule, the main component of the dentin organic matrix, consists of 2 α1 and one α2 chain intertwined in a triple helix, which contains a rigid helical center with N and C globular terminals called telopeptides (Figure 1A). These individual strands are composed of a coding system formed by the presence of specific Gly-XY amino acid sequences where commonly Gly, X and Y are glycine, proline and hydroxyproline, respectively, which ensure that each area of the individual α chains can bind to 4 adjacent helical molecules, forming a structure that resembles a propeller called microfibril [13,14] (Figure 1B).
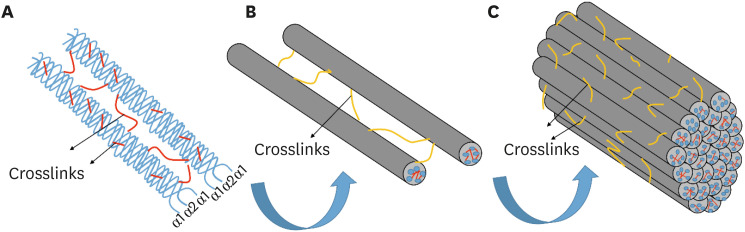 | Figure 1Structural organization of type I collagen fibril. (A) Two α1 and one α2 chains intertwine in a triple helix, giving rise to a collagen molecule. (B) Five adjacent helical molecules join together to form a microfibril. (C) Microfibrils meet in concentric bundles known as collagen fibrils. The crosslinking agents reinforce this bonding through the formation of crosslinks (represented in red and yellow) at the molecular and fibrillar levels, improving the mechanical properties of the demineralized collagen matrix.
|
The molecules are staggered in a way that a pattern of repetition of hundreds of molecules is created every 67 nm, typically known as the periodic D bandage of type I collagen. Staggered spaces between the ends of the successive collagen molecules produce the so-called gap zones, and areas, where multiple molecules are superimposed, represent the overlap zone, forming peaks and valleys and giving rise to the surface nano-structural roughness of microfibrils [15,16]. This repeated pattern may interconnect so that each valley (gap zone) is adapted to the peaks (overlap zone) of an adjacent microfibril. Thus, microfibrils meet in concentric bundles, better known as collagen fibrils, measuring about 100 nm in diameter [17] (Figure 1C).
Collagen fibrils, mainly type I, make up 30% of the volume of the hybrid layer that is the basis for a wide variety of restorative procedures currently performed in dentistry. The ultimate goal of adhesive procedures is the infiltration of monomers from adhesive systems and complete coverage of demineralized collagen fibrils. Thus, the network of collagen fibrils acts as an anchor to the resin, allowing the retention of adhesive restorations to the dentin [6,18].
However, in regions where the infiltration of the monomers is less than the depth of dentin demineralization resulting from the application of acidic substances, the formation of void spaces occurs where the fibrils remain unprotected. Therefore, they are susceptible to the action of organic matrix proteases, which promote the slow but continuous degradation of the collagen fibrils that make up the hybrid layer, leading to loss of bond strength [2,19].
MMPs are present in dentin and are known to degrade almost any component of the extracellular matrix, including collagen. Therefore, they have been considered to play a major role in the degradation of bare collagen fibrils within the hybrid layer [20].
Metalloproteinases: structure and mechanism of action
MMPs are a family of structurally related but genetically distinct enzymes that can degrade components of the extracellular matrix [21]. Twenty-four types of MMPs have already been identified (Table 1) and, because it was believed that each MMP had a specific substrate, the nomenclature was based on substrate specificity and molecular structure; thus the MMPs were classified as collagenases (MMP-1, MMP-8, MMP-13 and MMP-18), gelatinases (MMP-2 and MMP-9), stromelysins (MMP-3, MMP-10 and MMP-11), matrilysins (MMP-7 and MMP-26), membrane type MMP (MMP-14, MMP-15, MMP-16, MMP-17, MMP-24 and MMP-25), and others (MMP-12, MMP-19, MMP-20, MMP-21, MMP-23, MMP-27 and MMP-28). However, it is now recognized that MMPs generally degrade multiple substrates, but the substrate-based classification is still widely used [22].
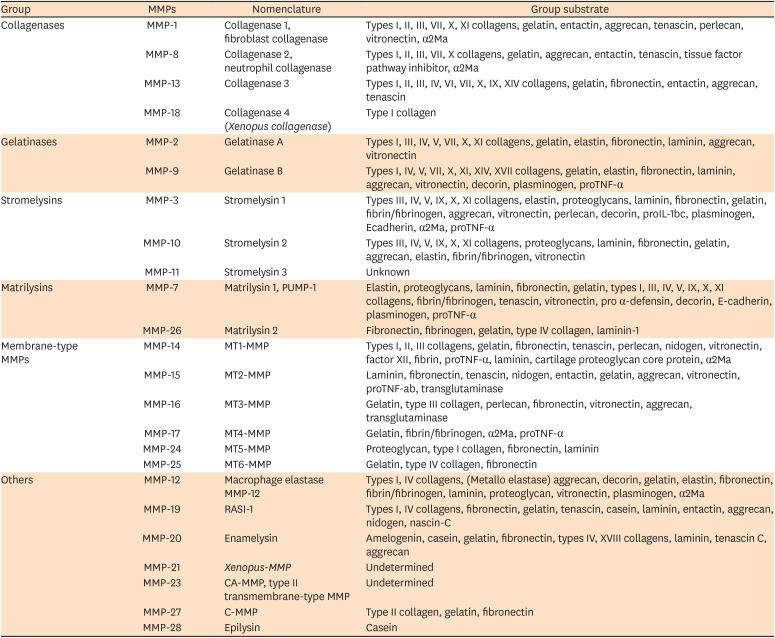
Table 1
Matrix metalloproteinases (MMPs) description, nomenclature and their substrate (modified from Hannas et al. [22])
MMPs based on substrate specificity and molecular structure. Groups of MMPs are listed with their trivial names.
![]()
Structurally, these enzymes are divided into several domains, which differ in dependence on each subtype. However, most MMPs are composed of an N-terminal propeptide, a catalytic domain, a hinge region and a C-terminal hemopexin domain [23]. The N-terminal propeptide contains approximately 80 amino acids, guaranteeing the latency of the enzyme. The most important functional amino acid within the N-terminal propeptide is cysteine, which interacts with zinc ions (Zn2+) in the catalytic site through a thiol group and constitutes the cysteine switch. The catalytic domain contains 2 Zn2+ and 2 or 3 calcium ions (Ca2+). The first Zn2+ present in the active site is directly involved in catalytic processes. The second Zn2+ so-called structural Zn2+ and Ca2+, which are approximately 12 nm away from the Zn2+ at the catalytic site, are required to stabilize the structure of that domain. The C-terminal domain is structurally similar to proteins in the hemopexin family and regulates the activity and specificity of the catalytic domain function. The catalytic domain and the C-terminal are freely bound by a flexible proline-rich peptide bond (hinge region). The length of the hinge region is extremely variable and the specific function is not fully understood [24].
MMP-2 and -9 also contain a fibronectin-like domain that is responsible for the recognition of collagen so-called collagen-binding domain [25]. Collagen binding domains are the primary sites of interaction with collagen and gelatin; they also mediate elastin binding. Therefore, they are required for the cleavage of collagen and elastin [26].
In the human body, MMPs are not only involved in negative actions but also play a significant protective role in human physiology. MMPs are important as regulators of postnatal development and remodeling and repair of tissues in response to injury, infection or disease [27]. In the oral environment, these enzymes are related to periodontal diseases, caries progression, pulpal inflammation and cancer. They may also be involved in the release of dentinal growth factors forming tertiary dentin in response to external aggression, tissue destruction in response to pulpal inflammation and, more recently, in the demineralized collagen decomposition of the hybrid layer of composite resin restorations [28,29].
Zymographic analyses have demonstrated that MMP-2, MMP-8, MMP-9, MMP-13 and MMP-20 are present in the dentin organic matrix. Of these, MMP-2 and MMP-9 are the most prevalent forms in dentin; MMP-2 is present in larger amounts than other forms of MMPs in human dentin [30,31].
In the mineralized matrix, the presence of apatite crystals on collagen fibrils and their non-collagenous proteins probably block the access of the catalytic site of the collagen, thus leaving the inactive MMPs. However, the acidic environment generated by adhesive procedures and caries processes removes the apatite crystals that cover the collagen and disrupts cysteine-zinc coordination, which is then lost; as a consequence of this, activation of latent proteases occurs [9,32,33] (Figure 2A). Although a small fraction of these proteases can be extracted by acids, most remain bound to the matrix in their active form, where they can slowly hydrolyze the collagen matrix [34] and represent a threatening to the integrity of the dentin/resin interface resulting on reduced durability of the adhesive restorations over time.
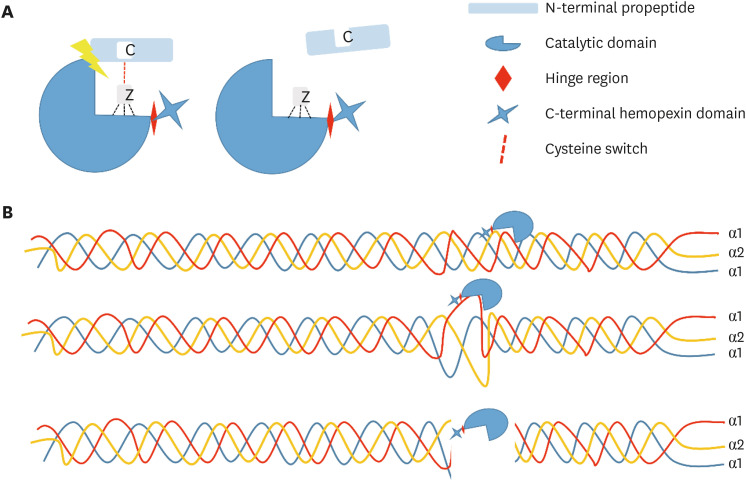 | Figure 2Structure and activation of metalloproteinases. (A) Metalloproteinases are composed of an N-terminal propeptide, catalytic domain, hinge region and C-terminal hemopexin domain. Its activation occurs when cysteine-zinc coordination is lost as a consequence of the activation of the proteases that were in latent form (proenzymes) in the tissue. (B) After activation, collagenase cleaves the collagen molecules, causing them to lose the triple helical conformation, resulting in 1/4 and 3/4 length fragments that will be slowly hydrolysed by the gelatinases.
|
Collagenase cleaves the triple helical collagen molecules into 2 fragments of one-quarter and 3-quarters length; the first quarter contains the C-terminal part of the collagen molecule and the second 3-quarters contain the N-terminal portion [35] (Figure 2B). Collagen loses its triple helical conformation and can then be degraded by gelatinases MMP-2 and MMP-9 [21]. MMP-2 and -9 attack the non-helical fragments and reduce them to even smaller peptide fragments. The binding sites of collagen to MMPs are very similar to the catalytic site of these enzymes. Thus, the collagen-binding domain of MMP-2 and -9 binds to a short segment of the collagen α1 chain [36]. After binding to the substrate, the water molecule that is bound to the zinc of the catalytic domain attacks the substrate and, through the transfer of protons, promotes the cleavage of the peptides [25]. It is speculated that endogenous MMPs in the dentin matrix bind to the collagen peptides via their hemopexin-like domains (MMP-1, -8) and/or their fibronectin-like domains (MMP-2, -9) [34].
Clinically, the degradation of the adhesive interface over time causes marginal discoloration, secondary caries and subsequent loss of retention. As a consequence, it is necessary to replace the restorations within a relatively short time. Therefore, in order to achieve an effective and stable bond, the preservation of dentin collagen is critical, because collagen represents the main organic component of the dentin matrix and plays a key role in bond strength [37,38].
Strategies to improve the stability of the dentin/resin bonding
In dentistry, there is concern about the long-term stability of the adhesive interface [22]. Synthetic or natural bioactive agents, which inhibit the endogenous enzymes of the organic matrix of collagen, have been used in the attempt to produce a strong and durable adhesive interface. The use of these substances may help to preserve the hybrid layer and increase the durability of the resin-tooth bond [39]. These agents can be applied in different ways: incorporated in the acid for dentin etching [40,41], included into the composition of the adhesive system [42-45] or applied as a solution directly on the dentin surface after the acid treatment remaining in contact with the surface of the dentin [46-49]. MMPs inhibitors act by different mechanisms such as by chelating cations [50-52], collagen crosslinking [53-55] and competitive inhibition for active sites of the collagen molecule [26,56,57].
1. Inhibition by chelating
Most of these agents act by inhibiting MMP activity by chelating Zn2+ from the active site [50]. Chlorhexidine (CHX) is the most well-documented MMP inhibitor. It has been shown that when applied after acid etching or incorporated into adhesive systems, it can preserve the integrity of the demineralized collagen matrix and increase the strength of the resin-dentin bond over time [58,59]. This substance acts as a non-specific inhibitor of MMPs, since it is capable of altering the three-dimensional structure of these enzymes and chelating the metal ions (Ca2+, Zn2+), which are necessary for the activation of its enzymatic function [56]. CHX is able to inactivate all MMPs in dentin even at low concentrations such as 0.02% [60]. Since the mechanism of CHX binding to dentin is only electrostatic [61], there is concern that CHX may have been displaced by cations from dentinal fluid or saliva loosening their anti-proteolytic feature. This may explain recent observations of da Silva et al. [58] and Montagner et al. [49] that showed CHX maintains the stability of the resin-dentin bond only up to 12 months. Therefore, the time of action of CHX as an inhibitor of MMPs is a limiting factor for its use [49,51]. To overcome this issue, de Menezes et al. [43] added a CHX modified release system using clay as a reinforcing agent and release modulator to an adhesive. CHX + clay showed better MMPs inhibition capacity than CHX and maintained the bone strength even after 18 months.
Some studies have used CHX diacetate instead of CHX digluconate because it preserved the resin-dentin bonding and did not interfere with water sorption, solubility or conversion degree of adhesive systems [58,62]. The main difference between these 2 chemicals is that CHX digluconate is only available as an aqueous solution while CHX diacetate is available as a powder. The addition of CHX digluconate to adhesives means adding water to the formulation, which may spoil the properties of the adhesive system. A new approach was attempted introducing a novel drug-delivery approach to demineralized dentin substrates and the resin-dentin interface with Poly(ε-caprolactone) CHX-loaded nanocapsules (nano-PCL/CHX). The authors showed infiltration retention of nano-PCL/CHX (125:25 and 125:50) inside dentinal tubules at depths of approximately 5 µm to10 µm [63].
In vivo, the short-term results are encouraging. Ricci et al. [64] and Brackett et al. [48] showed a beneficial effect of CHX on slow down the rate of resin-dentin bond degradation in the first months of function. But after 2 years, clinical trials showed that the addition of CHX into adhesive systems or its use as dentin pretreatment did not add clinical advantages and did not improve the clinical durability of adhesive restorations [65-68].
Similar to CHX, ethylenediaminetetraacetic acid (EDTA) and tetracyclines act by non-specific chelation of Zn2+ and Ca2+ cations [51]. Tetracyclines have been shown to inhibit collagen degradation and bone loss in periodontitis. The binding of calcium and zinc to the carbons -11 and -12 of tetracycline seems to be involved in this mechanism of inhibition [69], while EDTA contains 4 carboxylic acid groups and at neutral pH, it acts as a chelating agent [70].
Galardin, a synthetic MMP inhibitor, acts specifically by Zn2+ chelation located in the catalytic domain, selectively inhibiting MMP-2, -3, -8 and -9, in contrast to the mechanism of action of CHX. This inhibitor has a structure similar to collagen, which facilitates its binding to the active site of MMPs. Galardin inhibits the proteolytic activity in the demineralized dentin in a similar way to CHX for a period of 12 months, even when used in a concentration of approximately 10 to 100 times lower (0.2 mM) for 30 seconds. This result supports the concept that specific inhibitors act at very low concentrations [52]. Therefore, the use of galardin as an inhibitor of dentin MMPs may be a clinically relevant alternative to improve the stability of the hybrid layer. However, researchers still are searching for products that keep its effect lasts longer.
Quaternary ammonium compounds (QACs) are a group of cationic antimicrobial agents that can bind to negatively charged phosphate and carboxylic groups in hydroxyapatite and collagen, respectively, exhibiting similar anti-MMP action to CHX [71]. However, the nature of these electrostatic interactions is weak and similarly to biguanides, QACs are water-soluble molecules and may leach out of adhesive interface, reducing their anti-proteolytic effect overtime [72].
Daood et al. [73] synthesized 2%, 5% and 10% quaternary ammonium silane (QAS) and showed that QAS inhibited endogenous MMPs and cysteine cathepsins. Later, Daood et al. [74] used QAS on etched dentin before the bonding procedure. The use of a tetrafunctional organosilane as an anchoring unit for the antimicrobial trialkoxysilane molecules enables a three-dimensional network to be formed once condensation has been completed within the dentinal substrate. This minimizes the possibility for QAS molecules to leach out of the resin-dentine interface and gives a considerably longer activity. The 2% QAS is a good inhibitor of dentin proteases and could be an alternative for 2% CHX as MMPs inhibitor to improve the durability of the resin-dentin bond [74].
Benzalkonium chloride (BAC) is an antimicrobial agent containing a quaternary ammonium group. It has demonstrated inhibitory potential against endogenous proteases [42]. It is commercially available as an etchant (phosphoric acid-containing BAC 1.0 wt%) with controversial results [40,72]. Sabatini et al. [72] showed the effectiveness of BAC in preventing the bonding interface breakdown after 6 months. Contrary, on El Gezawi et al. [40] study, BAC-etch did not improve bond stability after 6 months. The etchant possesses anti-proteolytic properties. However, rinsing off the etchant technique limits the BAC available and in consequence its effect. Comba et al. [42] added BAC into the primer and adhesive formulation to get a more effective delivery system. Although the BAC-doped adhesive showed inhibitory property on dentin MMPs at baseline, this activity was not reflected by the bond strength results after 6 months. In general, BAC should be extensively tested before translation to clinical use.
A monomer in the family of QACs 12-methacryloyloxy-dodecyl-pyridinium bromide is a chemical incorporated into commercially available adhesives and showed dentin MMP inhibition similar to CHX but required higher concentrations. The advantage over other MMPs inhibitors is that it can copolymerize with adhesive monomers. So, its effect on MMPs can be expected to maintain for years [71,72]. Long-term clinical studies are necessary to validate that assumption.
Alcohols can also act as MMPs inhibitors. As hydrolases, MMPs require water for their proteolytic activity. Specifically, they add water to specific peptide bonds breaking large peptide chains into smaller ones [30,75]. Water is present in adhesives and in CHX solution (as the vehicle for its delivery), which results in increased water content into the hybrid layer. Tezvergil-Mutluay et al. [75] have tested a series of alcohols (methanol, ethanol, propanol, butanol, pentanol, heptanol and octanol, the ethanol ester of methacrylic acid and propanediol). Alcohols were able to inhibit human recombinant MMP-9 and the endogenous proteases of dentin matrix in a dose-dependent manner. Based on the molar concentration required to inhibit rhMMP-9 50%, 2-hydroxyethyl methacrylate (HEMA), 3-hexanol, 3-heptanol and 1-octanol showed the strongest inhibition. The results indicated that alcohols with 4 methylene groups inhibit MMPs more effectively than methanol or ethanol.
The mechanism of MMP inhibition of these alcohols seems to involve hydrophobic forces since the inhibitory activity of alcohol is inversely related to their concentration. High alcohol concentrations may denature enzymes by removing water from the enzyme structure, causing their denaturation. The presence of bound water prevents interpeptide hydrogen bonding within the peptide chains of enzymes. Removal of that water allows rapid interpeptide hydrogen bonding that can disrupt the tertiary structure that is necessary for enzyme activities. Some proteins can be recovered when the alcohol is removed and the proteins are rehydrated. Other proteins are unable to reconstitute their original tertiary structure [75].
In this context, ethanol wet-bonding was introduced in order to address a solution for improving resin-dentin bond durability. This technique is based on applying ethanol 100% or in ascending concentrations (50%, 70%, 80%, 95% and 3 × 100%) on demineralized dentin [76]. It was proposed aiming to remove the remaining water, creating wider interfibrillar spaces that allow the monomers to infiltrate deeply through the collagen matrix. However, ethanol wet bonding causes collapse and shrinkage of collagen fibrils and consequently poor resin infiltration. In addition, ethanol is not so effective in inhibiting MMPs [76]. So, the immediate benefits of the ethanol-bonding technique observed in the laboratory setting were not confirmed when the same protocol was performed in vivo [76].
2. Collagen crosslinking
In addition to the use of metalloproteinases inhibitors, collagen crosslinking agents have been used in an attempt to improve the mechanical properties of the demineralized collagen matrix (Figure 1C). This group includes proanthocyanidin (PA) [53,77], glutaraldehyde [78], baicalein [55], hesperidin [45], riboflavin [54,79], resveratrol [46] and quercetin [46].
PA can simultaneously perform a protease inhibition function, in addition to the collagen crosslinking function. The crosslinking effect of PA is attributed to its interaction with proteins rich in proline, which promotes the formation of intermolecular crosslinks within the collagen matrix and acts to increase the mechanical stability of the matrix [53]. Although CHX can successfully prevent degradation of the collagen matrix, it has no effect on the reinforcement of demineralized collagen fibrils [80]. Liu et al. [77] showed that 2% PA applied on the demineralized dentin for 30 seconds was able to increase the collagen resistance to degradation by MMPs.
When added to 10% phosphoric acid, PA did not compromise adhesion efficiency in enamel and dentin, and produced a stable dentin bond after six months of aging in water. Stability of the resin/dentin interfaces produced with this acid can be attributed to several factors that function synergistically, such as the ability to interact with collagen even in an acidic environment; their ability to inactivate the catalytic site of proteases and the increased resistance of collagen fibrils that render them less susceptible to degradation by proteases [38]. However, the long-term performance of the experimental material was not observed in this study.
Glutaraldehyde has also been used with the goal of improving the properties of demineralized dentin. Its ability to irreversibly bind proteins and increase the modulus of elasticity of collagen fibrils is of great interest in enhancing the collagen structure. Moreover, its ability to react chemically with collagen and resin components, such as HEMA, can also contribute to the penetration of the adhesive system and facilitate the wettability of the dentin substrate [78]. However, glutaraldehyde has high cytotoxicity and therefore, the use of other collagen crosslinkers with no or low toxicity is highly desirable [81].
Baicalein is one of the major flavonoids in Scutellaria baicalensis which inhibits the activity of MMP-2 and MMP-9 via PI3K/AKT, P38-MAPK-NF-κB and MAPK-ERK1/2 signal pathways. Moreover, baicalein and PAs have a similar molecular structure with phenolic hydroxyl functional groups, suggesting that they have comparable crosslinking properties. Used as dentin preconditioner followed by an etch-and-rinse adhesive system, it effectively improves the resin-dentin bonding durability [55]. Hesperidin is also a flavonoid. It was tested as MMPs inhibitor by Islam et al. [45], and it was effective in preserve dentin collagen what favors tissue remineralization due to keeping the collagen scaffold.
Riboflavin is a suitable agent for the crosslinking of dentin collagen because of its biocompatibility and its ability to produce free radicals when photo-activated in a spectrum ranging from ultraviolet (UV) to visible light (270 nm, 366 nm and 445 nm). These free radicals, or so-called reactive oxygen species, such as O2 and O2−, are released when the riboflavin is photo-activated and light is absorbed, forming covalent crosslinks between the adjacent collagen molecules. Riboflavin enhances immediate bond strength to dentin, stabilizes the adhesive interface and inhibits dentin MMPs, enhancing the durability of resin-dentin bonds. Although the use of ultraviolet A (UVA) is effective for photo-activation of riboflavin for collagen crosslinking, safety issues regarding the use of UVA and its practicality for dental use should be considered. Conventional blue halogen light units may be a possible alternative to replace UVA light sources because of their ready availability and safe ease of use in dentistry [54,79]. However, further studies investigating the use of visible light sources, which act in wide amplitude between UV and blue light, with coverage of the different absorption peaks of riboflavin are recommended, in order to better evaluate the penetration of visible light and activation depths of the riboflavin in the dentin matrix and its interaction with MMPs and monomers [54].
3. Competitive inhibition for active sites of the collagen molecule
Another mechanism proposed in the attempt to inhibit the action of MMPs is the competitive inhibition through the binding to the active sites of the collagen molecule. It has been demonstrated that Zn2+ or other divalent metals may reduce the expression of some MMPs. Zn2+ can be considered a competitive inhibitor of MMPs, exerting a protective effect through binding to the collagen-sensitive cleavage sites of the metalloproteinases. It seems that subtle conformational changes occur in collagen after zinc binding and lead to the protection of metalloproteinases cleavage sites [56]. The concentration of Zn2+ around matrix-bound MMPs may be very important in determining their activity on dentin collagen [56,82].
Osorio et al. [56] showed by evaluation of carboxyterminal telopeptide concentration of type I collagen (ICTP) released during degradation of collagen showed that the use of an adhesive loaded with zinc oxide (ZnO) nanoparticles reduced and stabilized the degradation of collagen for up to 4 weeks without negative interfering in dentin bond strength. However, it is crucial to analyze the ability of nanoparticles to release ions in a sustained manner and to determine the right concentrations of Zn2+ to be used without causing toxicity [83]. Since zinc oxide-containing materials have been widely used in the dentin interface over time, it is expected that new adhesive systems with zinc oxide will also be compatible, making their use viable.
Santamaria et al. [84] reported a series of 4,4'-biphenylsulfonamides as selective inhibitors of MMP-2 and -13 and showed that these molecules do not bind the catalytic zinc of MMPs. The authors suggest that this class of molecules binds to a catalytic subsite, possibly the S1'-pocket. All the compounds had a bent 4-substituted phenyl ring linked by 1C bridge to the 4'-position of a 1,1'-biphenyl-4-sulfonamide scaffold and, in general, resembled the structure of some previously reported MMP-13 non-zinc binding inhibitors. Two of the characterized inhibitors (11 and 19) act as non-competitive inhibitors, whereas the para-methyl ester derivative 13 behaves as a competitive inhibitor.
The study conducted by Xu et al. [26] identified a collagen peptide, which can bind specifically to the collagen-binding domain of MMP-2. Binding of the peptide to this domain blocks the positioning of substrate molecules relative to the catalytic site and thereby inhibits the activities of MMP-2.
4. Inhibitors with multiple action mechanisms
Carbodiimides have been used as an alternative to glutaraldehyde because they do not contain potentially cytotoxic aldehyde residues. It inhibits MMPs by inactivating the active sites by reducing their molecular mobility and also improves the resistance of cross-linked collagen matrices to degradation by inducing exogenous crosslinks and thereby increasing their stiffness. Their non-specifically crosslink proteins, activating the carboxylic acid groups of glutamic and aspartic acids to form an O-acylisourea intermediate. The latter reacts with the ε-amino groups of lysine or hydroxylysine to form amide crosslinking, leaving the urea as the terminal by-product. Carbodiimides are an example of crosslinking agents in which the peptides are crosslinked to one another without introducing additional linking groups. Thus, this crosslinking agent increases the mechanical properties of the collagen matrix and thus renders resin-dentin bonds more durable [85]. In addition, this substance is a potent inhibitor of MMPs. Its inhibitory effect, due to reduced molecular mobility, is much faster than the crosslinking effect of collagen [86]. Therefore, it seems that the inactivation of MMP by substances such as carbodiimide will have a longer-lasting effect than the exclusively MMP inhibiting substances, possibly due to the combined action of these 2 mechanisms of action that favor the maintenance of the hybrid layer integrity.
Resveratrol and quercetin as polyphenols can inhibit MMP activity by zinc chelation [12]. They have a high complexation affinity to metal ions, and zinc is essential for MMP activity [87]. Quercetin itself increases the expressions of tissue inhibitor of metalloproteinase (TIMP-2) [88]. Resveratrol and quercetin suppress prostaglandin-E synthesis by inhibiting the activity of cyclooxygenase-2 enzyme (COX-2) and also inhibit COX-2 gene transcription. It may be particularly useful because the over-production of the COX-2 induces MMP release. Gotti et al. [89] showed that a total-etch 2-step quercetin-doped adhesive (5.0 w%) significantly increased the bond strength after 180 days. Porto et al. [46] tested quercetin and resveratrol as pretreatment of etched dentin. Comparisons between 1 day and 120 days microtensile bond strength measurements showed that after 120 days the results were similar to or higher than CHX.
Green tea has also been proposed as a natural inhibitor of MMPs. This inhibitory action has been attributed mainly to polyphenols in the tea, especially epigallocatechin gallate [90]. This polyphenol is capable of stabilizing the collagen chain by increasing the number of crosslinks of the collagen fibrils and reducing biodegradation of collagen. The mechanism of action of epigallocatechin gallate is complex, involving multifunctional activities such as downregulation of endogenous protease expression, inactivation of protease and prevention of free access of collagenase to sites active in collagen chains [39].
Monteiro et al. [90] showed that although the application of the green tea solution reduced bond strength after 24 hours, compared to the control. Contrary, after 6 months, the union strength values did not present statistically significant differences compared to the control groups and CHX. In another study, when aqueous extract of epigallocatechin gallate was applied for a shorter time than green tea solution, there was no negative interference in the immediate bond strength and the stability of the adhesive interface was maintained (6 and 12 months) similar to CHX [39]. Probably the negative results observed in the short-term evaluation (24 hours) in the study conducted by Monteiro et al. [90] could have been due to the interference of other components of green tea that were not present in the extract of epigallocatechin gallate isolated from green tea.
The inhibition of MMPs has been shown to reduce the degradation of the adhesive interface when applied directly to the dentin as a pre-treatment. However, this strategy increases the clinical steps of adhesive application [9]. When substances were incorporated into components of the adhesive systems, concerns such as substantivity issues, which restricted the time of action of these inhibitors and their release from polymerized resin matrices, which could limit their inhibitory potential [91]. Thus, results obtained with these strategies are not linear, showing neutral, positive or negative effects on bonding stability [58], most likely due to the different techniques of application and the wide variety of substances used for that. Thus, more studies should be conducted to better clarify the details about controlling the action of MMPs, reinforcement collagen by the crosslinking agents, as well as long-term tests to observe how long the protective effect against MMPs will last, and thus targeting the results to identify ideal conditions for its use that support its clinical indication. From future research, new types of adhesive systems could be developed and are likely to be available in the near future, improving the quality of dentin adhesion techniques, providing a more stable adhesion, capable of resisting collagenolytic degradation, resulting in restorative procedures with higher quality and longevity.
Clinical remarks and perspectives on further researches
Data from studies included in this review have demonstrated that endogenous enzymes contained in the dentin are involved in the failure of adhesive restorations [48,58,59,64]. Slowing down or preventing the irreversible destruction of the teeth organic matrix would allow obtaining long-lasting restorations. A promising overview of MMPs inhibitors activity coming from in vitro studies had been found in this review. However, a lack of long-term clinical trials was noticed. Table 2 resumes the researches on exogenous MMP inhibitors, their mechanism of action, type of study, and period of the test.
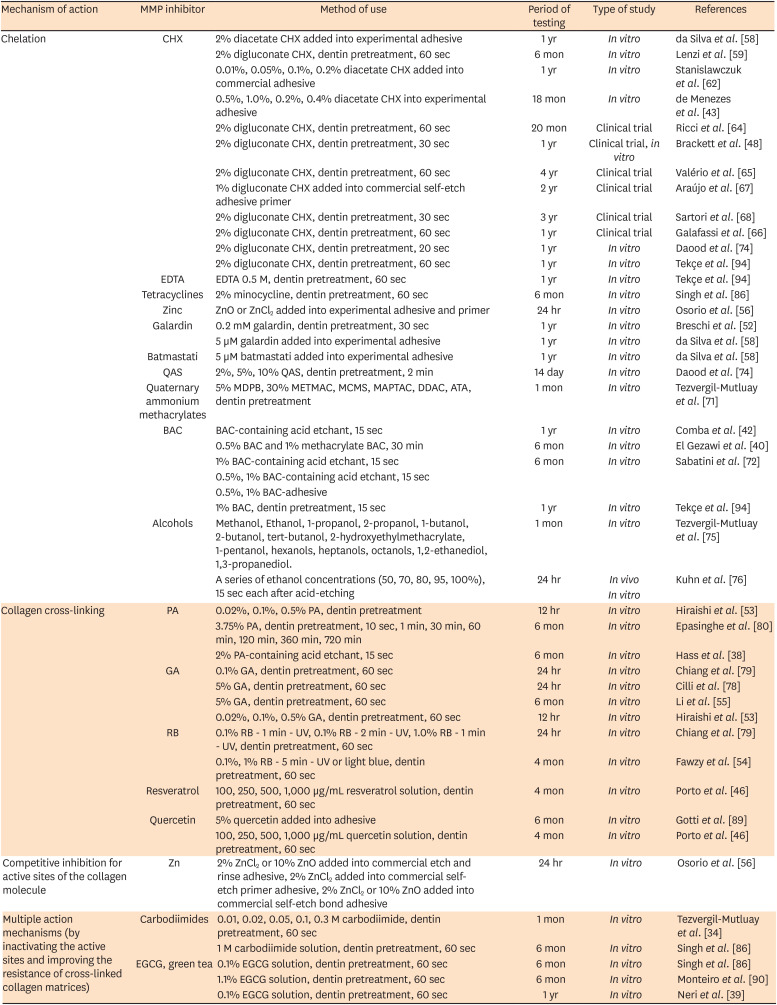
Table 2
Matrix metalloproteinases (MMPs) inhibitors and strategies to improve the stability of the dentin/resin bonding
| Mechanism of action | MMP inhibitor | Method of use | Period of testing | Type of study | References |
|---|---|---|---|---|---|
| Chelation | CHX | 2% diacetate CHX added into experimental adhesive | 1 yr | In vitro | da Silva et al. [58] |
| 2% digluconate CHX, dentin pretreatment, 60 sec | 6 mon | In vitro | Lenzi et al. [59] | ||
| 0.01%, 0.05%, 0.1%, 0.2% diacetate CHX added into commercial adhesive | 1 yr | In vitro | Stanislawczuk et al. [62] | ||
| 0.5%, 1.0%, 0.2%, 0.4% diacetate CHX into experimental adhesive | 18 mon | In vitro | de Menezes et al. [43] | ||
| 2% digluconate CHX, dentin pretreatment, 60 sec | 20 mon | Clinical trial | Ricci et al. [64] | ||
| 2% digluconate CHX, dentin pretreatment, 30 sec | 1 yr | Clinical trial, in vitro | Brackett et al. [48] | ||
| 2% digluconate CHX, dentin pretreatment, 60 sec | 4 yr | Clinical trial | Valério et al. [65] | ||
| 1% digluconate CHX added into commercial self-etch adhesive primer | 2 yr | Clinical trial | Araújo et al. [67] | ||
| 2% digluconate CHX, dentin pretreatment, 30 sec | 3 yr | Clinical trial | Sartori et al. [68] | ||
| 2% digluconate CHX, dentin pretreatment, 60 sec | 1 yr | Clinical trial | Galafassi et al. [66] | ||
| 2% digluconate CHX, dentin pretreatment, 20 sec | 1 yr | In vitro | Daood et al. [74] | ||
| 2% digluconate CHX, dentin pretreatment, 60 sec | 1 yr | In vitro | Tekçe et al. [94] | ||
| EDTA | EDTA 0.5 M, dentin pretreatment, 60 sec | 1 yr | In vitro | Tekçe et al. [94] | |
| Tetracyclines | 2% minocycline, dentin pretreatment, 60 sec | 6 mon | In vitro | Singh et al. [86] | |
| Zinc | ZnO or ZnCl2 added into experimental adhesive and primer | 24 hr | In vitro | Osorio et al. [56] | |
| Galardin | 0.2 mM galardin, dentin pretreatment, 30 sec | 1 yr | In vitro | Breschi et al. [52] | |
| 5 µM galardin added into experimental adhesive | 1 yr | In vitro | da Silva et al. [58] | ||
| Batmastati | 5 µM batmastati added into experimental adhesive | 1 yr | In vitro | da Silva et al. [58] | |
| QAS | 2%, 5%, 10% QAS, dentin pretreatment, 2 min | 14 day | In vitro | Daood et al. [74] | |
| Quaternary ammonium methacrylates | 5% MDPB, 30% METMAC, MCMS, MAPTAC, DDAC, ATA, dentin pretreatment | 1 mon | In vitro | Tezvergil-Mutluay et al. [71] | |
| BAC | BAC-containing acid etchant, 15 sec | 1 yr | In vitro | Comba et al. [42] | |
| 0.5% BAC and 1% methacrylate BAC, 30 min | 6 mon | In vitro | El Gezawi et al. [40] | ||
| 1% BAC-containing acid etchant, 15 sec | 6 mon | In vitro | Sabatini et al. [72] | ||
| 0.5%, 1% BAC-containing acid etchant, 15 sec | |||||
| 0.5%, 1% BAC-adhesive | |||||
| 1% BAC, dentin pretreatment, 15 sec | 1 yr | In vitro | Tekçe et al. [94] | ||
| Alcohols | Methanol, Ethanol, 1-propanol, 2-propanol, 1-butanol, 2-butanol, tert-butanol, 2-hydroxyethylmethacrylate, 1-pentanol, hexanols, heptanols, octanols, 1,2-ethanediol, 1,3-propanediol. | 1 mon | In vitro | Tezvergil-Mutluay et al. [75] | |
| A series of ethanol concentrations (50, 70, 80, 95, 100%), 15 sec each after acid-etching | 24 hr | In vivo | Kuhn et al. [76] | ||
| In vitro | |||||
| Collagen cross-linking | PA | 0.02%, 0.1%, 0.5% PA, dentin pretreatment | 12 hr | In vitro | Hiraishi et al. [53] |
| 3.75% PA, dentin pretreatment, 10 sec, 1 min, 30 min, 60 min, 120 min, 360 min, 720 min | 6 mon | In vitro | Epasinghe et al. [80] | ||
| 2% PA-containing acid etchant, 15 sec | 6 mon | In vitro | Hass et al. [38] | ||
| GA | 0.1% GA, dentin pretreatment, 60 sec | 24 hr | In vitro | Chiang et al. [79] | |
| 5% GA, dentin pretreatment, 60 sec | 24 hr | In vitro | Cilli et al. [78] | ||
| 5% GA, dentin pretreatment, 60 sec | 6 mon | In vitro | Li et al. [55] | ||
| 0.02%, 0.1%, 0.5% GA, dentin pretreatment, 60 sec | 12 hr | In vitro | Hiraishi et al. [53] | ||
| RB | 0.1% RB - 1 min - UV, 0.1% RB - 2 min - UV, 1.0% RB - 1 min - UV, dentin pretreatment, 60 sec | 24 hr | In vitro | Chiang et al. [79] | |
| 0.1%, 1% RB - 5 min - UV or light blue, dentin pretreatment, 60 sec | 4 mon | In vitro | Fawzy et al. [54] | ||
| Resveratrol | 100, 250, 500, 1,000 µg/mL resveratrol solution, dentin pretreatment, 60 sec | 4 mon | In vitro | Porto et al. [46] | |
| Quercetin | 5% quercetin added into adhesive | 6 mon | In vitro | Gotti et al. [89] | |
| 100, 250, 500, 1,000 µg/mL quercetin solution, dentin pretreatment, 60 sec | 4 mon | In vitro | Porto et al. [46] | ||
| Competitive inhibition for active sites of the collagen molecule | Zn | 2% ZnCl2 or 10% ZnO added into commercial etch and rinse adhesive, 2% ZnCl2 added into commercial self-etch primer adhesive, 2% ZnCl2 or 10% ZnO added into commercial self-etch bond adhesive | 24 hr | In vitro | Osorio et al. [56] |
| Multiple action mechanisms (by inactivating the active sites and improving the resistance of cross-linked collagen matrices) | Carbodiimides | 0.01, 0.02, 0.05, 0.1, 0.3 M carbodiimide, dentin pretreatment, 60 sec | 1 mon | In vitro | Tezvergil-Mutluay et al. [34] |
| 1 M carbodiimide solution, dentin pretreatment, 60 sec | 6 mon | In vitro | Singh et al. [86] | ||
| EGCG, green tea | 0.1% EGCG solution, dentin pretreatment, 60 sec | 6 mon | In vitro | Singh et al. [86] | |
| 1.1% EGCG solution, dentin pretreatment, 60 sec | 6 mon | In vitro | Monteiro et al. [90] | ||
| 0.1% EGCG solution, dentin pretreatment, 60 sec | 1 yr | In vitro | Neri et al. [39] |
CHX, chlorhexidine; EDTA, ethylenediaminetetraacetic acid; QAS, quaternary ammonium silane; METMAC, [2(methacryloyloxy)ethyl] trimethylammonium chloride; MCMS, methacryloyl choline methyl sulfate; MAPTAC, [3(methacryloylamino)propyl] trimethylammonium chlorid; DDAC, diallyldimethylammonium chloride; ATA, 2-acryloxyethyltrimethylammonium chloride; MDPB, 12-methacryloyloxydodecylpyridinium bromide; BAC, benzalkonium chloride; PA, proanthocyanidin; GA, glutaraldehyde; RB, riboflavin; Zn, zinc; EGCG, epigallocatechin-3-gallate.
![]()
New approaches have attempted to address some of the drawbacks of classic MMP inhibitors such as nanocarriers for drug delivery [43]. Chemicals were added to commercial and experimental adhesives [42-44] and researches have been carried out with methacrylamide-based monomers as novel materials for use in dental adhesives, hydrolytically stable, showing increased resistance to enzymatic degradation [92,93].
Most of the preclinical studies on exogenous MMP inhibitors still are in the early stages and some of them showed limited success. Moreover, the great difficulty in translating laboratory into clinical researches is the study design regarding the selection of concentrations, drug-releasing, and period of the test. The lack of standardized methodologies makes rough the evaluation of its efficacy.
New MMP inhibitors with higher selectivity and substantivity could improve the specificity of MMP inhibitors targeting specific MMPs involved in dentin proteolysis. Besides, because of the differences in the proteolytic activities of different MMPs towards different substrates, it is critical to examine different MMP inhibitors in various regions of dentin and at different stages of the caries disease. Urge to investigate its ability against MMPs action through more in vivo research with extended follow-up.
Go to :
CONCLUSION
Selective inhibitors of MMPs promise to be the future for specific targeting of preventing dentin proteolysis. The knowledge about the MMPs functionality should be considered to synthesize drugs capable to efficiently and selectively block the MMPs chemical routes targeting their inactivation in order to overcome the current limitations of the therapeutic use of MMPs inhibitors, i.e., easy clinical application and long-lasting effect.
Go to :
 XML Download
XML Download