

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Melanin is produced by melanocytes located in the basal layer of the epidermis to protect skin cells against harmful external agents, including ultraviolet (UV) rays, fine dust, and various chemical compounds [1]. The factors secreted from keratinocytes mainly by UV irradiation, including alpha-melanocyte-stimulating hormone (α-MSH), adrenocorticotropic hormone, stem cell factor, endothelin 1, hepatocyte growth factor, basic fibroblast growth factor, and granulocyte-macrophage colony-stimulating factor, stimulate each receptors on the surface of melanocytes and melanin is produced in lysosome-related organelle called melanosome [2-7]. Among these factors, α-MSH is a small peptide hormone derived from pro-opiomelanocortin. Binding of α-MSH to melanocortin 1 receptor increases cyclic adenosine monophosphate (cAMP) level that activates protein kinase A (PKA) [8,9]. PKA then phosphorylates cAMP responsive element binding protein (CREB) and transcription of microphthalmia-associated transcription factor (MITF) is induced by phospho-CREB [10]. MITF acts as a transcription factor for tyrosinase, tyrosinase-related protein 1 (TYRP1), and dopachrome tautomerase (Dct or TYRP2), key enzymes in melanogenesis, and induces their expression [11]. The matured melanosome moves toward the dendrite tip, and melanosome motor protein complex consisting of Rab27a, Melanophilin, and Myosin Va plays a role in this process [12-14]. Melanophilin, which has binding domains for Rab27a, Myosin Va, and actin, simultaneously binds to Rab27a on the melanosome surface and to Myosin Va interacting with actin filament. The assembly of these motor proteins is required for melanosome transport to the dendrite tips along actin filament [15-19]. Subsequently, melanosomes transferred from dendrite tips of melanocytes to neighboring keratinocytes and incorporated melanosomes locate in the perinuclear area of keratinocytes [20,21]. Although proper melanin formation is important for normal cell protection, excessive melanin production induces hyperpigmentation, including melasma, freckles, and solar lentigo, which can lead to mental stress and poor quality of life [22,23]. Thus, there are demands for the development of satisfactory whitening agents without side effects such as skin irritation, allergy induction, and carcinogenesis and many studies targeting various whitening mechanisms have been conducted [24,25].
Peptides are short chain of amino acids, the smallest unit of protein, linked to each other by peptide bonds. Recently, extensive research and development regarding peptides as active ingredients have been performed in the cosmetics and pharmaceutical industries because of their high biocompatibility and protein mimicking ability [26,27].
We screened our peptide library and found four peptides having each effect on inhibition of melanin synthesis, migration and transfer as well as melanosome degradation promoting activity. Subsequently, this study aimed to investigate the whitening activity of a peptide mixture containing four peptides with the same molar ratio.
METHODS
Materials
The 2-CTC resin used in the peptide synthesis was purchased from BeadTech (Seoul, Korea). Fmoc-amino acid, hydroxybenzotriazole (HOBt), and N,N,N´,N´-tetramethyl-O-(1H-benzotriazol-1-yl)uronium hexafluorophosphate (HBTU) were purchased from CS Bio Co. (San Francisco, CA, USA). The reagents used in the synthesis, including dimethylformamide (DMF), N,N-diisopropylethylamine (DIEA), piperidine, trifluoroacetic acid (TFA), thioanisole, phenol, ethane dithiol (EDT), triisopropylsilane (TIS), and diethyl ether, were purchased from Daejung Chemical & Metal Co., Ltd. (Seoul, Korea).
Dulbecco's Modified Eagle Medium (DMEM) powder for melanocytes and fetal bovine serum (FBS) were purchased from Thermo Fisher Scientific (Waltham, MA, USA), and liquid DMEM media and penicillin/streptomycin were purchased from Welgene Inc. (Gyeongsan, Korea). Sodium bicarbonate, sodium hydroxide, arbutin, 3-[4,5-dimethylthiazole-2-yl]-2,5-diphenyltetrazolium bromide (MTT), 3,4-dihydroxy-L-phenylalanine (L-DOPA), trypsin, and rapamycin were purchased from Sigma-Aldrich (St. Louis, MO, USA), and Solvable was purchased from PerkinElmer (Waltham, MA, USA). Antibodies against Rab27A, Melanophilin, MITF, Tyrosinase, and Actin were purchased from Santa Cruz Biotechnology (Dallas, TX, USA), and antibodies against Beclin-1, LC3, p62, p-CREB and p-ERK1/2 were purchased from Cell Signaling Technology (Danvers, MA, USA). Primer synthesis was performed in Cosmo GENETECH (Seoul, Korea) and further used in the experiment (Table 1).
Peptide synthesis
CTC resin with a capacity of 0.84 mmol/g was subjected to swelling reaction in a reactor containing DMF solvent. Then, 2 equivalents of Fmoc-C terminal amino acid and 2.5 equivalents of DIEA were added in DMF to the reactor and further subjected to reaction for 2 h. The termination of the reaction was confirmed with a Kaiser test kit (Sigma-Aldrich), and the resin was washed with DMF. The Fmoc was removed by adding 20% piperidine/DMF twice to the washed resin. The termination of the reaction was confirmed with the Kaiser test kit, and the resin was washed with DMF. Subsequently, the following process was repeated according to the sequence of amino acids in the C terminal to N terminal direction. After 2 equivalents of Fmoc-amino acid, 2 equivalents of HBTU, 2 equivalents of HOBt, and 2.5 equivalents of DIEA were added in DMF, the reaction was performed for 2 h. The termination of the reaction was confirmed with the Kaiser test kit, and the resin was washed with DMF. The Fmoc was removed from amino acid by adding 20% piperidine/DMF twice to the washed resin. The termination of the reaction was confirmed with the Kaiser test kit, and the resin was washed with DMF. In case of Pep-3, Ferulic acid conjugation was performed through the following procedure. After 2 equivalents of Ferulic acid, 2 equivalents of HBTU, 2 equivalents of HOBt, and 2.5 equivalents of DIEA were added in DMF, the reaction was performed for 2 h. The termination of the reaction was confirmed with the Kaiser test kit, and the resin was washed with DMF. After the final synthesis was completed, a cleavage solution (TFA:H₂O:thioanisole:phenol:EDT:TIS = 81.5:5:5:5:2.5:1) was added to separate the peptide from the resin. The peptide was then precipitated using diethyl ether and washed five times. Subsequently, drying was performed to recover the final peptide product.
The purity of the synthesized peptide was identified by HPLC (Thermo Fisher Scientific/U-3000) analysis, and it was detected at UV 214 nm under the flow rate of 1 ml/min under mobile phase 0.1% TFA in water/0.1% TFA in acetonitrile using a C18 (Agilent, Pursuit XRs, 250 × 4.6 mm, 5 μm, 100Å) column.
To identify the molecular weight, LC-MS/MS (AB SCIEX, 3200 Q-trap) analysis was performed, and it was detected by MS/MS at the flow rate of 0.25 ml/min under mobile phase 0.1% formic acid in water/0.1% formic acid in acetonitrile gradient using a C18 (Agilent, Pursuit XRs, 100 × 2.0 mm, 5 μm, 100Å) column. The MS/MS analysis conditions included ESI Positive mode, Source/Gas: CUR = 20, CAD = High, IS = 5500, TEM = 350, GS1 = 50, GS2 = 50/Compound DP = 50–80, EP = 10, CE = 10–50, and CES = 1–10 (Table 2).
Cell culture
The B16F10 melanoma cells used in this experiment were purchased from ATCC (Manassas, VA, USA). B16F10 cells were incubated in DMEM media containing 10% heat-inactivated FBS, 1% penicillin/streptomycin, and 1.5 g/L sodium bicarbonate at 37°C and under the condition of 5% CO2.
HaCaT keratinocytes were purchased from CLS (Eppelheim, Germany). HaCaT cells were incubated in DMEM media containing 10% heat-inactivated FBS and 1% penicillin/streptomycin at 37°C and under the condition of 5% CO2.
Cell viability
B16F10 cells were seeded in 48-well plate at a density of 8 × 103 cells/well and incubated for 16 hours. They were then treated with various concentrations of peptide mixture in 2% FBS containing media for 3 days. After incubation, 20 μl of 4 mg/ml MTT was treated to each well for 4 hours and the media was removed. Subsequently, cells were treated with 500 μl of DMSO to dissolve formazan. Absorbance was measured at 570 nm using a spectrophotometer (Molecular Devices, San Jose, CA, USA).
HaCaT cells were seeded in 96-well plate at a density of 5 × 103 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in serum-free media for 3 days. Subsequently, MTT assay was performed in the same manner as described above.
Melanin content assay
B16F10 cells were seeded in 6-well plate at a density of 5 × 104 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in 2% FBS containing media for 3 days. To induce melanin production, 100 ng/ml α-MSH was co-treated and 200 μM arbutin was used as a positive control. The cells were collected and lysed by treating with 200 μl of 1 M NaOH and absorbance for lysate at 490 nm was measured with a spectrophotometer.
Tyrosinase activity test
B16F10 cells were seeded in 6-well plate at a density of 5 × 104 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in 2% FBS containing media for 3 days. To induce melanin production, 100 ng/ml α-MSH was co-treated and 200 μM arbutin was used as a positive control. Each well was treated with 200 μl of lysis buffer (Sigma-Aldrich) to collect the lysate. After quantitation of the protein using BCA kit (Thermo Fisher Scientific), 90 μg of protein from each group was incubated with 20 μl of 10 mM L-DOPA in 96-well plate for 30 min at 37°C. Absorbance for mixture at 475 nm was measured with a spectrophotometer.
Melanosome isolation
B16F10 cells were seeded in 6-well plate at a density of 5 × 104 cells/well and incubated for 16 h. Cells were treated with 100 ng/ml α-MSH for 72 h to induce melanin synthesis and harvested by trypsin/EDTA treatment. Cells were then washed with 10 ml of 0.25 M sucrose (in 10 mM HEPES buffer) and resuspended in 10 ml of 0.25 M sucrose solution. Subsequently, after freezing with liquid nitrogen, thawing was performed at 37°C, which was repeated 10 times. After centrifugation at 1,000 g for 10 min, the supernatant was collected and further centrifuged for 45 min at 20,630 g and 4°C. Subsequently, the pellet was subjected to resuspension with DPBS and stored at –80°C before use.
Melanosome uptake test
HaCaT cells were seeded in 6-well plate at a density of 1 × 105 cells/well and incubated for16 h. They were then treated with the peptide mixture in serum-free media for 1 h and further treated with isolated melanosomes for 40 h. The cells were lysed by treating with 200 μl of 1 M NaOH and absorbance for lysate at 490 nm was measured with a spectrophotometer. Furthermore, melanosome staining was performed using a Fontana-Masson Stain Kit (ScyTek Laboratories, Logan, UT, USA) and images of cells were detected by light microscope.
Melanosome degradation test
HaCaT cells were seeded in 6-well plate at a density of 1 × 105 cells/well and incubated for 16 h. They were pretreated with isolated melanosomes in serum-free media for 48 h and additionally treated with the peptide mixture for 72 h. One micromole of rapamycin was used as positive control. The cells were lysed by treating with 200 μl of 1 M NaOH and absorbance for lysate at 490 nm was measured with a spectrophotometer. Furthermore, melanosome staining was performed using a Fontana-Masson Stain Kit and images of cells were detected by light microscope.
Gene expression analysis (RT-PCR)
B16F10 cells were seeded in 6-well plate at a density of 5 × 104 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in 2% FBS containing media for 3 days. To induce melanin production, 100 ng/ml α-MSH was co-treated and 200 μM arbutin was used as a positive control. RNA was isolated from cells using RNA extraction kit (Qiagen, Germany), and cDNA synthesis was performed using RT-PCR premix (iNtRON Biotechnology, Seongnam, Korea). After preparation of reaction mixture with PCR premix (iNtRON Biotechnology) and primers for each gene, PCR was performed using PCR machine (Eppendorf, Germany). Subsequently, mRNA expression patterns were determined by agarose gel electrophoresis.
HaCaT cells were seeded in 6-well plate at a density of 3 × 105 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in the serum-free media for 1 h and additionally treated with 4U trypsin for 16 h. The cells were collected and RT-PCR was performed in the same manner as described above.
Protein expression analysis (Western blotting)
B16F10 cells were seeded in 6-well plate at a density of 5 × 104 cells/well and incubated for 1 day. They were then treated with various concentrations of peptide mixture in 2% FBS containing media for 3 days. To induce melanin production, 100 ng/ml α-MSH was co-treated and 200 μM arbutin was used as a positive control. The cells were lysed with lysis buffer and the protein was quantified using BCA kit. Twenty micrograms of protein were separated on 8% polyacrylamide gel and electrophoretically transferred to polyvinylidene fluoride membranes. Membranes were blocked in 5% w/v skim milk in PBS with 0.5% Tween 20 (PBS-T). Incubation with primary antibodies diluted in blocking solution was performed overnight at 4°C and was followed by washing with PBS-T. The appropriate secondary antibodies were diluted in blocking solutions and incubated with the membranes for 1 h at room temperature followed by washing with PBS-T. The membranes were visualized via Western detection reagent (Elpis Biotech, Daejeon, Korea) with Gel Doc (Bio-Rad, Hercules, CA, USA).
For analysis of p-CREB and p-ERK, B16F10 cells were seeded in 6-well plate at a density of 3 × 105 cells/well and incubated for 1 day. They were then treated with various concentrations of peptide mixture in 2% FBS containing media. The incubation times were 30 min for analysis of p-CREB and 10 min for analysis of p-ERK. To induce melanin production, 100 ng/ml α-MSH was co-treated and 200 μM arbutin was used as a positive control. Western blotting was performed using isolated proteins in the same manner as described above.
HaCaT cells were seeded in 6-well plate at a density of 3 × 105 cells/well and incubated for 16 h. They were then treated with various concentrations of peptide mixture in serum-free media for 3 days. Two hundred nanograms of rapamycin was treated for 3 hours as a positive control. Western blotting was performed using isolated proteins in the same manner as described above.
Skin equivalent analysis
To test the whitening effect of peptide mixture using skin equivalent, liposome sample was prepared as follows. The peptide mixture was dissolved in distilled water at concentration of 2,000 μg/ml and adjusted to pH 7. After filtration using 0.22-μm pore size filter, 1% hydrogenated phosphatidylcholine buffer was mixed at a ratio of 10%. The nano-sized liposome sample was prepared by passing through the 75-μm cells five times at 1,000 bar using the Microfluidizer.
MelanoDerm MEL-300-B (MatTek Corporation, Ashland, MA, USA) was topically treated with 25 μl or 50 μl of liposome containing 2,000 μg/ml peptide mixture three times a week for 2 weeks. Same volume of vehicle was applied to control tissue. The treated skin equivalents were washed with DPBS and light microscopy was performed to check morphology of melanocytes. Subsequently, to analysis the production of melanin, skin equivalents were frozen at –80°C for 30 min and thawed at room temperature. Tissue samples were incubated with 1% sodium bicarbonate for 30 min and dried. After treatment with 300 μl of Solvable, the samples were left overnight at 95°C and then cooled at room temperature. Absorbance of the tissue extract at 490 nm was measured with a spectrophotometer.
To observe the melanin distribution within the tissues through histology, all samples were fixed in 4% phosphate-buffered formalin for 24 h. Fixed samples were paraffin-embedded and cut into a 5-μm section using microtome (Leica Biosystems, Wetzlar, Germany). Paraffin sections were stained using Fontana-Masson Stain Kit according to the manufacturer's instructions and light microscopy were performed.
Statistics
The experiments conducted in this study were repeated three times or more. The statistical significance of the data was tested through Student's t-test. The resulting values were expressed as mean ± standard deviation and were considered statistically significant when the p value was less than 0.05.
RESULTS
Peptide mixture inhibits α-MSH-induced melanin synthesis and tyrosinase activity in B16F10 cells
To determine the effect of the peptide mixture on cell viability, B16F10 melanoma cells and HaCaT keratinocytes were treated with the peptide mixture at concentrations of 10, 50, 100, and 200 μM, and MTT assay was performed. The results showed that peptide mixture was not cytotoxic in the overall treatment concentrations, and subsequent experiments were performed in the above concentration range (Fig. 1A).
To identify the inhibitory effect of the peptide mixture on melanin synthesis, B16F10 cells were treated at concentrations of 10, 50, 100, and 200 μM to compare melanin production rates. The results showed dose-dependent inhibition of melanin production by the peptide mixture. Inhibition rates were 3% at the lowest concentration of 10 μM and 26% at the highest concentration of 200 μM (Fig. 1B).
Tyrosinase is one of the key enzymes in melanin synthesis. The tyrosinase activity assay was performed to verify that the melanogenesis inhibitory effect of peptide mixture was caused by the regulation of tyrosinase. The results showed significant inhibition of tyrosinase activity by the peptide mixture in B16F10 cells. Inhibition rates were 8% at the lowest concentration of 10 μM and 33% at the highest concentration of 200 μM (Fig. 1C).
Peptide mixture inhibits the expression of melanosome biogenesis/transportation-related factors
To investigate the mechanism of depigmenting action shown by peptide mixture, expression levels of melanogenesis-related factors and melanosome transportation-related factors were analyzed.
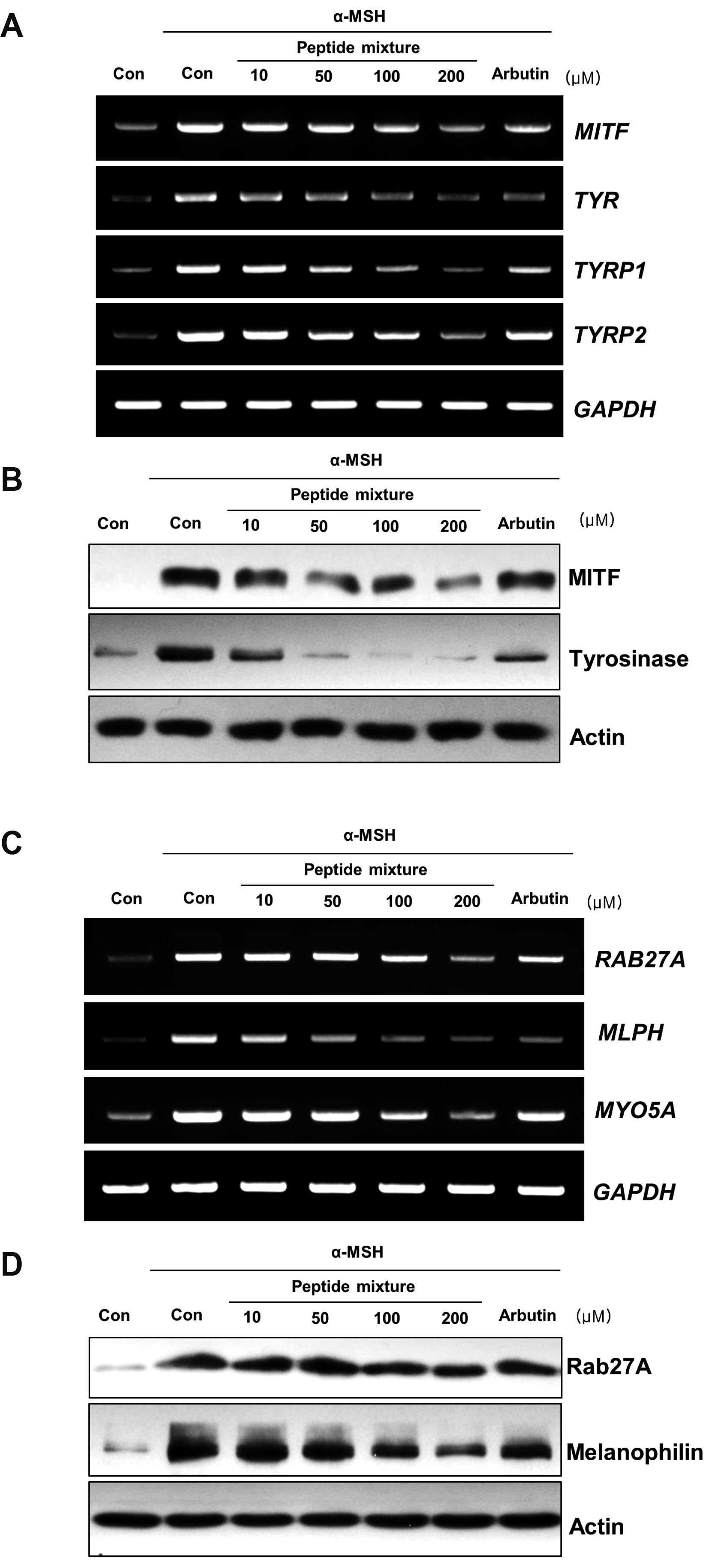
As shown in Fig. 2A, transcription levels of MITF and of its downstream genes, TYR, TYPR1 and TYRP2, were decreased by peptide mixture in B16F10 cells. Furthermore, α-MSH-induced protein levels of MITF and Tyrosinase were significantly decreased by peptide mixture in B16F10 cells (Fig. 2B).
These results suggest that the melanogenesis inhibition effect of peptide mixture occurred by inhibiting the expression of melanogenesis-related factors.
It has been published that MITF also acts as a transcription factor that activates motor proteins which have role in melanosome transportation [28,29]. To identify whether the peptide mixture, which has inhibitory effect on MITF expression, decreased the expression of motor proteins, we analyzed mRNA and protein levels. As shown in Fig. 2C, transcription levels of RAB27A, MLPH, and MYO5A genes were significantly decreased by peptide mixture in B16F10 cells. Furthermore, α-MSH-induced protein levels of Rab27a and Melanophilin were decreased by peptide mixture in B16F10 cells (Fig. 2D).
These results suggest that inhibition of MITF expression by peptide mixture can be result in inhibition of melanosome transportation.
Peptide mixture inhibits MITF activity through the regulation of CREB and ERK phosphorylation
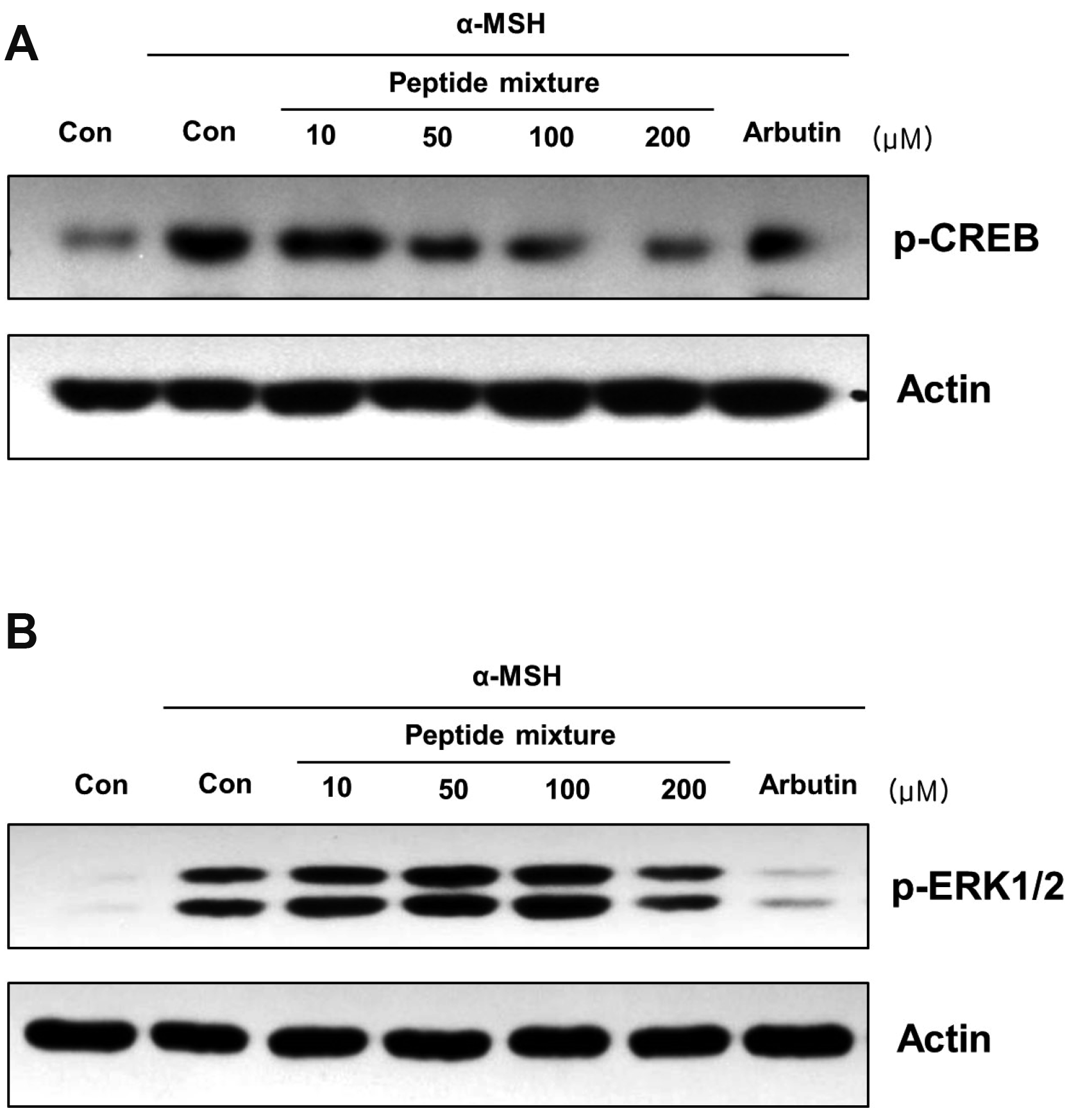
To investigate the action mechanism of peptide mixture on MITF expression, we analyzed phosphorylation level of transcriptional and post-translational regulatory factors of MITF.
As shown in Fig. 3A, phosphorylated CREB, a transcription factor activating MITF expression, was dose-dependently decreased by peptide mixture. Previously, it has been identified that activity of MITF depends on its post-translational modifications and proteasome-mediated MITF protein degradation can be induced by phosphorylated ERK1/2 [30]. Peptide mixture significantly increased p-ERK1/2 level under α-MSH-stimulated condition in B16F10 cells (Fig. 3B).
These results suggest that the expression and activity of MITF might be decreased through the regulation of CREB and ERK1/2 phosphorylations by peptide mixture.
Peptide mixture suppresses melanosome uptake of HaCaT keratinocytes
Melanosomes are known to be transferred from dendrite tips of melanocytes to keratinocytes through the phagocytosis of keratinocytes. To identify whether the peptide mixture could inhibit the melanosome transfer to keratinocytes, HaCaT keratinocytes were treated with the peptide mixture at concentrations of 10, 50, 100, and 200 μM and further treated with melanosomes isolated from B16F10 cells. The amount of melanosomes transferred to the cells was dose-dependently decreased by peptide mixture (Fig. 4A). Image analysis through Fontana-Masson staining showed that the distribution of brown-stained melanosomes significantly decreased in the peptide mixture-treated group in a dose-dependent manner (Fig. 4B).
Furthermore, the expression of F2RL1 (PAR-2) gene, a receptor that regulates the phagocytosis of keratinocytes, was significantly decreased by peptide mixture under trypsin-stimulated condition (Fig. 4C).
These results suggest that the peptide mixture can reduce phagocytosis through the inhibition of PAR-2 expression in keratinocytes by the peptide mixture.
Peptide mixture induces melanosome degradation in HaCaT keratinocytes
Melanosomes transferred into keratinocytes are known to be gradually eliminated by the cornification of keratinocytes or degraded by intracellular autophagy system. To identify whether the peptide mixture could promote the melanosome degradation, we treated peptide mixture to the melanosome containing HaCaT keratinocytes. When the amounts of melanosomes remaining was monitored, dose-dependent decrease was observed from the cell lysate treated with peptide mixture (Fig. 5A). Image analysis through Fontana-Masson staining showed that the distribution of brown-stained melanosomes significantly decreased in the peptide mixture-treated group in a dose-dependent manner (Fig. 5B).
Furthermore, when the autophagic activity related-protein levels in keratinocytes were monitored, the protein levels of Beclin-1 and LC3-II in relation to autophagosome formation were increased by peptide mixture treatment, while the protein level of p62 degraded by autophagolysosomes was decreased by peptide mixture treatment (Fig. 5C).
These results suggest that the peptide mixture induced the autophagic activity to promote the degradation of the melanosomes in keratinocytes.
Peptide mixture showed anti-pigmentation effect in skin equivalent model
An experiment using MelanoDerm was conducted to identify whether the peptide mixture could result in anti-pigmentation of the skin. After topical treatment of liposome containing 2,000 ppm of the peptide mixture was conducted at doses of 50 and 100 μg for 2 weeks, light microscopy was performed. The formation of dark melanosomes was induced entirely in the melanocyte bodies and dendrite tips of the control tissue, while the overall cell bodies were bright with a smaller number of dendrites because melanin production was inhibited in the tissue treated with the peptide mixture (Fig. 6A). Also, the melanin contents were dose-dependently decreased by peptide mixture, supporting the visual observation (Fig. 6B).
Moreover, histology was performed to observe the melanin distribution throughout the epidermis. According to the result, the control tissue showed a marked increase of melanogenesis in melanocytes of the basal layer, and brown-stained melanin was observed in keratinocytes due to the accompanied transfer to nearby keratinocytes. A considerable level of melanin was also observed in the upper cornified layers. On the other hand, the melanin contents of melanocytes were low in the tissues treated with the peptide mixture, and a significant decrease in melanin contents were observed throughout keratinocytes and cornified layers in comparison to the control group (Fig. 6C).
These results confirmed a significant anti-pigmentation effect of the peptide mixture on the skin equivalents.
DISCUSSION
The development of whitening materials has been targeted mainly to inhibit melanogenesis in melanocytes through the regulation of melanogenesis-related factors [31-35]. Recently, other targets of depigmentation, including melanosome migration, transfer and degradation, have been studied and several agents have been identified [36-42]. Because pigmentation is an action involving multiple mechanism, it might be efficient to use the combination of therapeutic agents which have regulatory effects for each target.
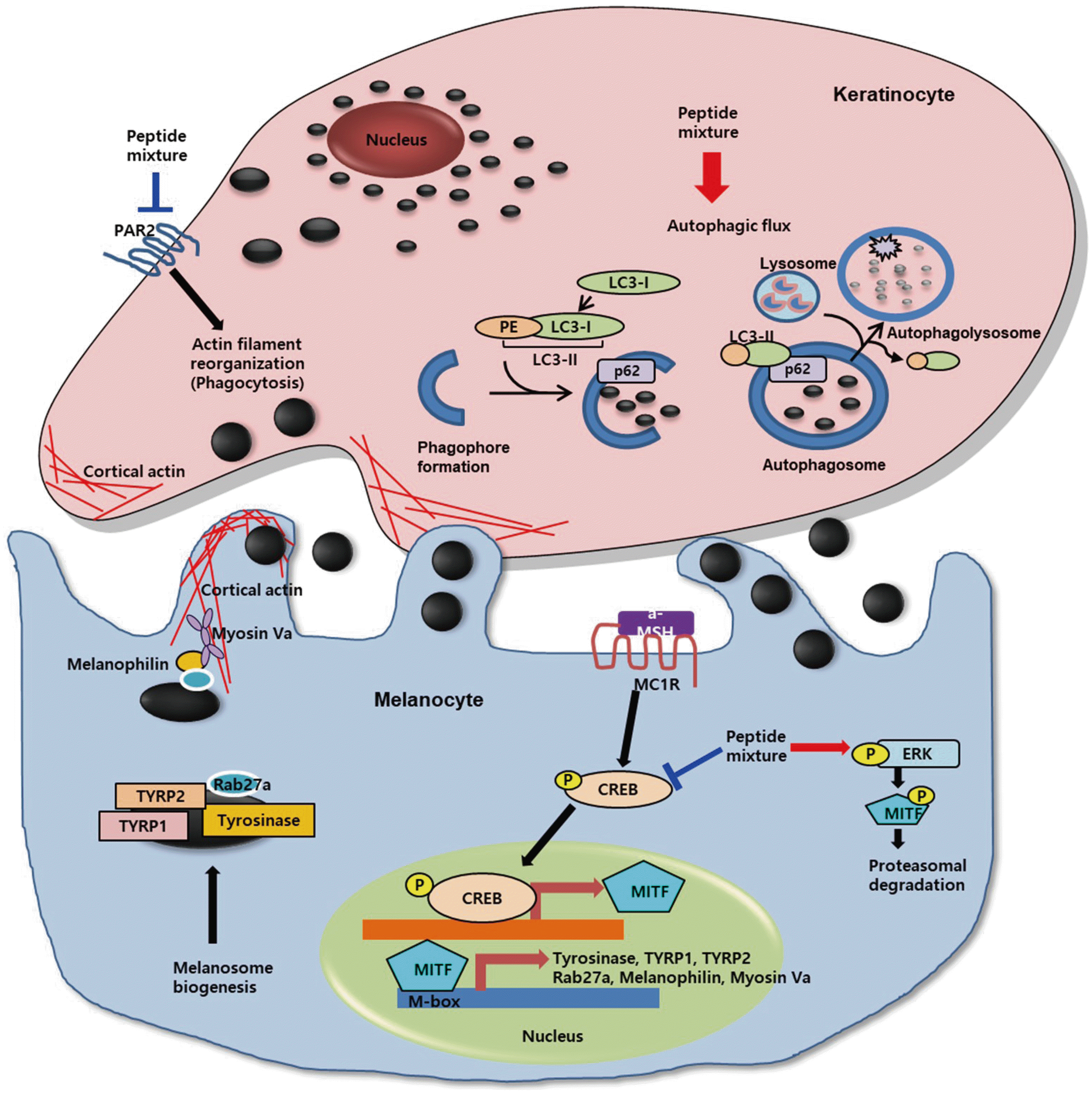
This study investigated the whitening activity of a peptide mixture containing four different peptides with the same molar ratio. The suggested action mechanism of peptide mixture against melanosome biogenesis, transfer and degradation is illustrated in Fig. 7. In melanocytes, the peptide mixture inhibits CREB phosphorylation results in decreased MITF expression and induces phosphorylation of ERK1/2 results in MITF phosphorylation and proteasomal degradation. As a result, the expression of melanosome biogenesis and transport-related proteins, including Tyrosinase, TYRP1, TYRP2, Rab27A, Melanophilin and MYO5A, are decreased. In keratinocytes, the peptide mixture decreases expression of PAR-2 that mediates keratinocyte phagocytic capability correlated with actin filament reorganization. Furthermore, the peptide mixture activates autophagic flux, and transferred melanosomes are degraded in autophagolysosome.
Peptides have been extensively examined as active ingredients for cosmetics because of their high biocompatibility and protein-mimicking activity [43]. But they also have some disadvantages that they can be easily degraded by proteases present in the skin and cannot easily penetrate the epidermis composed of lipids due to their hydrophilicity [44]. The studies for increasing peptide stability have suggested the methods of substituting amino acids at predicted cleavage site, acetylation of the N terminal, and amidation of the C terminal [45]. Furthermore, there are many studies to improve the transdermal penetration of peptides: methods involving the use of penetration inducers such as alcohols, azones, hexanoates, and unsaturated fatty acids; methods involving the use of specific peptide sequences that facilitate permeation; methods combining lipophilic derivatives; and methods encapsulating peptides such as liposomes, transfersomes, niosomes, and ethosomes [46-48]. In this regard, we prepared liposome encapsulating the peptide mixture to test its whitening effect in skin equivalent model by topical treatment, and significant reduction of melanin contents throughout the epidermal layers was shown in the test liposome treatment group (Fig. 6C).
In conclusion, the peptide mixture of this study exhibited whitening effect through various actions, including inhibition of melanin synthesis and migration as well as inhibition of melanosome uptake to keratinocytes and promotion of melanosome degradation. The peptide mixture of this study could be used as a new whitening material and that the liposome that facilitates the stability and skin penetration of peptides could be used to develop effective whitening products.


 XML Download
XML Download