

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Cerebral cortex integrates bottom-up sensory and top-down associative information in a multilayered structure consisting of excitatory pyramidal neurons (PyNs) with intermingled inhibitory interneurons. PyNs in layers 2/3 (L2/3) and 5 receive the two types of information mainly in basal dendrites and distal apical dendrites, respectively [1]. Multiple basal dendrites of L2/3 PyNs surround the soma and a single apical dendrite is extended into layer 1 (L1) as dendritic tufts [2]. These two dendritic compartments exhibit different properties of synaptic transmission and integration due to differential expression of ion channels and intracellular signaling molecules [3]. Neuromodulators such as acetylcholine and serotonin exert a profound effect on a vast array of neuronal functions in the central nervous system, such as neuronal excitability, learning and memory, experience-dependent development and brain state [4,5]. Since neuromodulators control information flow depending on brain state [6], the layer-specific neuromodulation of synaptic transmission might be critical to brain state-dependent functions. In our previous studies, muscarinic stimulation induced long-term potentiation (LTP) or endocannabinoid-dependent long-term depression (LTD) in basal dendrites, but not in distal apical dendrites of L2/3 PyNs in the primary visual cortex [7,8]. Serotonin (5-HT) facilitates GABAergic currents in perisomatic area, but not in distal apical dendrites of the same PyNs [9]. However, this differential neuromodulation between two dendritic compartments is not known in other cortical areas such as prefrontal association cortex.
The prefrontal cortex (PFC) is critical for high-level executive functions and cognitive control by integrating the processed sensory and emotional signals [10-13]. Among the prefrontal brain regions, the orbitofrontal cortex (OFC) including the agranular insular (AI) cortex plays an essential role in value-based decision making [14-16]. In the OFC, the feedforward inputs from other cortex and inputs from the amygdala terminate preferentially onto L2/3 while nonspecific thalamic relay nucleus and higher order cortex projects mainly onto L1 in rats [17-19] and macaque monkeys [20]. Especially, the thalamic nucleus submedius projects primarily to layers 2/3/5 of the OFC in mice [21]. The PFC also receives dense serotonergic innervation from dorsal and medial raphe nuclei and cholinergic innervation from the basal forebrain and basal ganglia [22,23]. The gating and polarity of synaptic plasticity in the cortex can be controlled by neuromodulators such as 5-HT and acetylcholine (ACh) [24-26]. The regulatory roles of various serotonergic and cholinergic stimulation in the induction of LTP and LTD of the medial PFC (mPFC) are relatively well established [25,27,28]. However, long-term synaptic plasticity in the lateral portion of the PFC such as AI cortex is not known. Moreover, the layer-specific modulation of synaptic transmission in L2/3 PyNs would be critical for the cognitive function of the PFC.
We investigated the layer-specific neuromodulation of synaptic transmission as a general phenomenon in the neocortex by analyzing the layer-specific serotonergic and cholinergic induction of long-term synaptic plasticity of glutamatergic transmission in L2/3 PyNs of the OFC. We found that serotonin induced input-specific LTD on perisomatic area, but not in distal apical dendrites. In contrast, cholinergic stimulation induced muscarinic LTD in both inputs. Our findings suggest that layer-specific neuromodulation of synaptic transmission might be generally applicable to the neocortex, and thus presents a novel cellular mechanism in cognitive processes.
Go to : 
METHODS
Slice preparation
Coronal slices of the frontal cortex were prepared from Sprague-Dawley (SD) rats (Orient Bio Co., Ltd., Seongnam, Korea) on days 21–27 after birth, which were raised under standard conditions (23°C ± 1°C, 12/12 h/ light/dark cycle). Animal care and surgical procedures were conducted by the institutional animal care and use committee (IACUC) of the Catholic University of Korea (IACUC approval No. 2019-0024-01) and were consistent with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. Animals were sedated with chloral hydrate (400 mg/kg, i.p.). Brains were rapidly isolated after decapitation and immersed in ice-cold artificial cerebrospinal fluid (ACSF) containing (in mM) 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 1 CaCl2, 2 MgSO4, and 10 D-glucose bubbled with 95% O2 and 5% CO2. Coronal slices (300-µm thick) were prepared on a vibratome (Microm HM650V, Walldorf, Germany) and incubated in a submerged slice chamber for 30 min at 36.5°C, and then left at room temperature (22°C–24°C) until the recording was performed. Slices were transferred to the recording chamber and continuously perfused with the same ACSF (bubbling with 95% O2 and 5% CO2) except for the addition of 2 mM CaCl2 and 1 mM MgSO4 (1–1.5 ml/min).
Whole-cell patch-clamp recording
The whole-cell patch-clamp recording was performed as described previously [7]. Recording electrodes (4–6 MΩ) were pulled from borosilicate glass pipettes (World Precision Instruments, Inc., Sarasota, FL, USA) using a micropipette puller (MODEL P-97; Sutter Instrument Co., Novato, CA, USA). Excitatory postsynaptic potential (EPSP) was recorded with an electrode filled with a solution containing (in mM) 130 K-gluconate, 10 KCl, 4 Mg-ATP, 10 Na2-phosphocreatine, 0.3 Na3-GTP, and 10 HEPES (pH 7.25 with KOH). The shape of L2/3 PyNs (~300–350 µm from pia) in the AI area of the PFC (Paxinos and Watson [29]) was visually identified with an infrared-differential interference contrast (IR-DIC) video microscope coupled to an upright microscope (BX50-WI fitted with 40x/0.80 NA immersion objective; Olympus, Tokyo, Japan) and their regular spiking patterns were confirmed. Whole-cell recording signals were acquired using an amplifier (IX2-700; Dagan, Minneapolis, MN, USA) and digitized at 20 kHz (Digidata 1322A; Axon Instruments, Foster City, CA, USA). Using pCLAMP 10.3 software (Axon Instruments), the signals were saved to a computer hard drive (Pentium PC). The data were low-pass filtered at 5 kHz. Membrane potentials were not corrected for ~14 mV calculated junction potential. The typical access resistance was 12–15 MΩ, and the bridge balance set was optimized in the experiment. The input resistance was measured by injecting negative current (–50 pA, 150 ms) into the soma (every 60 sec). Bipolar tungsten electrode (100 µm in diameter, SNE-100; Kopf Instruments, Tujunga, CA, USA) and glass electrode (20-µm tip diameter, filled with ACSF) were located at layer 3 (L3, ~50 µm downward and ~100 µm laterally from the soma) and corresponding L1 (~50 µm from pia), respectively, for the local electrical stimulations. EPSPs were evoked by paired-pulse stimulation (40-ms interval) with a brief square current pulse (0.2-ms duration) using a constant-current isolation unit (A360; World Precision Instruments). The EPSP amplitude changes were monitored at 0.05 Hz. The stimulus intensity was adjusted to obtain stable baseline EPSPs (5–8 mV). The average amplitude of EPSP was measured at 30–40 min after wash-out of serotonin or carbachol (CCh) and compared with that of baseline EPSPs. In some experimental sets, 6,7-dinitroquinoxaline-2,3-dione disodium salt (DNQX, 20 µM) and d-(-)-2-amino-5-phosphonopentanoic acid (D-AP5, 50 µM) were added to ACSF to confirm glutamatergic synaptic transmission. The paired-pulse ratio (PPR) was determined as the amplitude of the second EPSP compared with that of the first EPSP evoked paired-pulse stimulation (40-ms interval). The amplitude of the second EPSP was determined from the single exponential fit of the first EPSP.
FM1-43 unloading
The unloading protocol of fluorescence dye, FM1-43 was similar to that described previously [8]. The changes in fluorescence intensity of FM1-43 indicate synaptic vesicle recycling at presynaptic terminals. Synaptic stimulation releases the FM1-43 from presynaptic vesicles resulting in decreased fluorescence intensity of FM1-43 [30]. To load FM1-43 (Molecular Probes, Eugene, OR, USA) into synaptic vesicles, slices were exposed to high-potassium (40 mM) ACSF for 2 min with 8 µM FM1-43. To reduce nonspecific binding of the FM1-43 to tissue, slices were washed with 1 mM ADVASEP-7 for 2 min in ACSF and further perfused with 0.1 mM ADVASEP-7 in ACSF for 50 min. To unload FM1-43, electric stimulation (2 Hz) was delivered to either L1 or L3 for 2 min with glass pipettes or tungsten bipolar electrodes with 2- to 3-fold stimulus intensity for baseline EPSP amplitude. All loading and unloading procedures of FM1-43 were conducted using ACSF containing ionotropic glutamatergic blockers D-AP5 (50 µM) and DNQX (20 µM) to prevent excitotoxic damage. High-potassium ACSF was applied for 5 min at the end of experiments for the background subtraction and normalization of fluorescence signal. Fluorescence signals were acquired on the microscope for whole-cell patch-clamp recording equipped with a monochromator (Polychrome V; TILL Photonics GmbH, Gräfelfing, Germany), a long-pass filter (510 nm), and a CCD camera (Retiga-2000RV; QImaging, Surrey, BC, Canada). Wavelengths for excitation and emission of FM1-43 dye were 480 nm and 532 nm, respectively. Fluorescence images were collected at 5-sec intervals with 10–70 ms light exposure and captured using TILLvisION software (TILL Photonics).
Drugs
Drugs including 5-Hydroxytryptamine (5-HT), DNQX, 8-hydroxy-2-dipropylaminotetralin hydrobromide (8-OH-DPAT), N-(piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide (AM251), D-AP5, 1-(m-chlorophenyl) biguanide hydrochloride (mCPBG), NAN-190, ketanserin, and 1,2-bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid (BAPTA) were purchased from Tocris Bioscience (Bristol, UK). ondansetron hydrochloride, α-methyl-5-hydroxytryptamine (α-me-5-HT), CCh, and atropine were purchased from Sigma-Aldrich (St Louis, MO, USA). BAPTA was included in the pipette solution. NAN-190 and AM251 were dissolved in dimethyl sulfoxide (DMSO, 0.1%).
Statistical analysis
Data are expressed as mean ± standard error. Statistical comparisons were performed using paired or unpaired two-tailed Student’s t-test and two-way ANOVA with Tukey’s multiple comparisons. The level of significance was set at p < 0.05.
Go to :
RESULTS
Layer-specific activation of layer 2/3 pyramidal neurons with local electrical stimulation
Whole-cell patch-clamp recording was performed in L2/3 PyNs (300–400 µm from the pia) of the AI cortex, a part of OFC [14,16], in 3-week-old rats with local electrical stimulation of corresponding L1 and L3. In the primary visual cortex, as reported previously, these local stimulations activated non-overlapping synapses located in distal apical dendrites and basal dendrites, respectively, using unloading experiment of FM1-43 dye [8]. In the present study, we analyzed the spatial profile of unloaded FM1-43 dye by stimulation of L1 or L3 in the OFC (Fig. 1A–C). Stimulation of L1 resulted in an apparent decrease in fluorescence almost limited to L1 (F8, 36 = 103.5, p < 0.001), and the stimulation was reduced far less in deeper L2/3 near the soma of PyNs. However, the repeated stimulation of L3 reduced fluorescence near the soma (300–600 µm from the pia). These results demonstrated that local electrical stimulation of L1 and L3 mainly activated synapses in distal apical dendrites and perisomatic dendrites, respectively. This distance profile of stimulation was similar to that of the primary visual cortex [8]. Based on these results, the two stimuli were used to evoke layer-specific activation of synaptic transmission in L2/3 PyNs of the OFC.
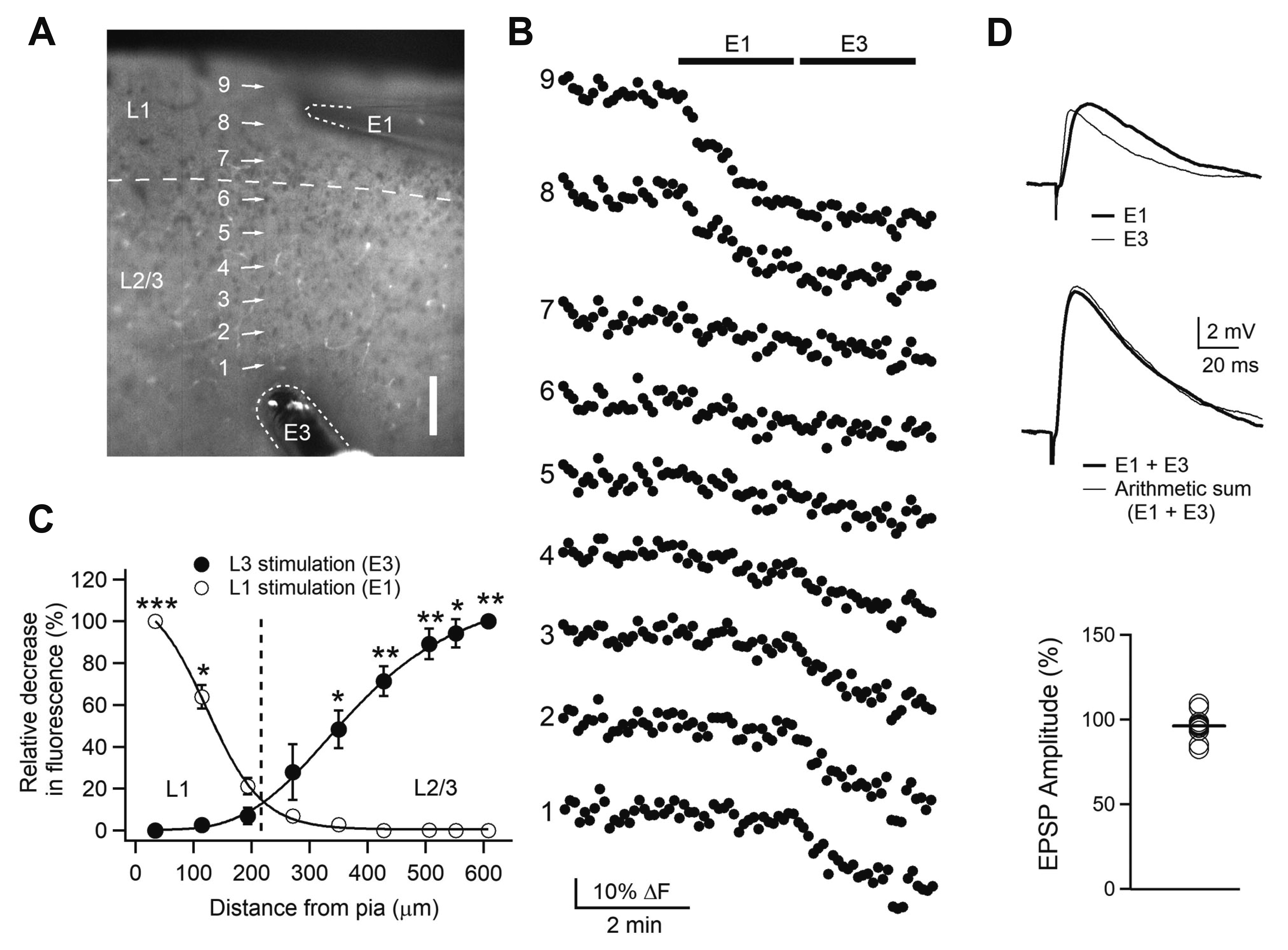 | Fig. 1Layer-specific activation of Layer 2/3 pyramidal neurons with local electrical stimulation.Experimental configuration (10× objective image) showing numbered regions of interest (ROIs) with stimulating electrodes marked with E1 for layer 1 (L1) stimulation and E3 for layer 3 (L3) stimulation. Scale = 100 μm. (B) Fluorescence changes in ROIs with time (5-sec interval). E1 and E3 were applied at 2 Hz (bars). Data are from a representative experiment (n = 3). (C) Spatial profiles of relative fluorescence changes against distance from the pia. The relative decrease in fluorescence to the maximal decrease was plotted. *p < 0.05, **p < 0.01, ***p < 0.001 compared with L1 or L3 stimulation. (D) Activation of non-overlapping synapses. Upper traces show representative EPSPs evoked by E1 and E3. Middle traces show representative EPSP evoked by combining E1 with E3 and arithmetic sum of corresponding E1-EPSP and E3-EPSP. Lower panel shows individual data of the amplitude of arithmetic sum relative to the amplitude of EPSPs evoked by combined E1 and E3 (n = 12). EPSP, excitatory postsynaptic potential.
|
Next, we investigated whether local electrical stimulation of L1 and L3 activated non-overlapping array of synapses (Fig. 1D). The amplitude of EPSPs evoked by simultaneous stimulation of L1 and L3 (9.5 ± 0.3 mV, n = 12) was similar to that of the arithmetic sum of each L1-EPSP and L3-EPSP (9.9 ± 0.4 mV, p = 0.107). The peak time (12.1 ± 1.2 ms) and decay time constant (58.8 ± 10.9 ms) of L1-EPSPs were longer than those of L3-EPSPs (6.4 ± 0.4 ms, p < 0.001 and 41.3 ± 5.2 ms, p < 0.05). Consistent with layer-specific activation of axon terminals demonstrated in the imaging experiment, these results indicate that the stimulation of L1 and L3 specifically activated synapses in distal apical dendrites and perisomatic dendrites including basal dendrites, respectively, in L2/3 PyNs of the OFC.
To investigate whether EPSP was mediated by AMPA- and NMDA-type glutamate receptors, we recorded EPSPs in the presence of the AMPA receptor blocker DNQX and the NMDA receptor blocker D-AP5 (Fig. 2). We evoked EPSPs with 5–8 mV amplitude in the present study. Application of D-AP5 (50 µM) did not change the amplitude of EPSPs (L1 stimulation: 100.6% ± 3.9%, n = 6, p = 0.877; L3 stimulation: 98.8% ± 1.3%, n = 5, p = 0.437). Co-application of DNQX (20 µM) with D-AP5 abolished synaptic response (L1 stimulation: 2.6% ± 0.6%, p < 0.001; L3 stimulation: 3.8% ± 0.4%, p < 0.001). These results indicate that EPSPs evoked by local electrical stimulation resulted mostly from the activation of AMPA receptors in L2/3 PyNs of the OFC.
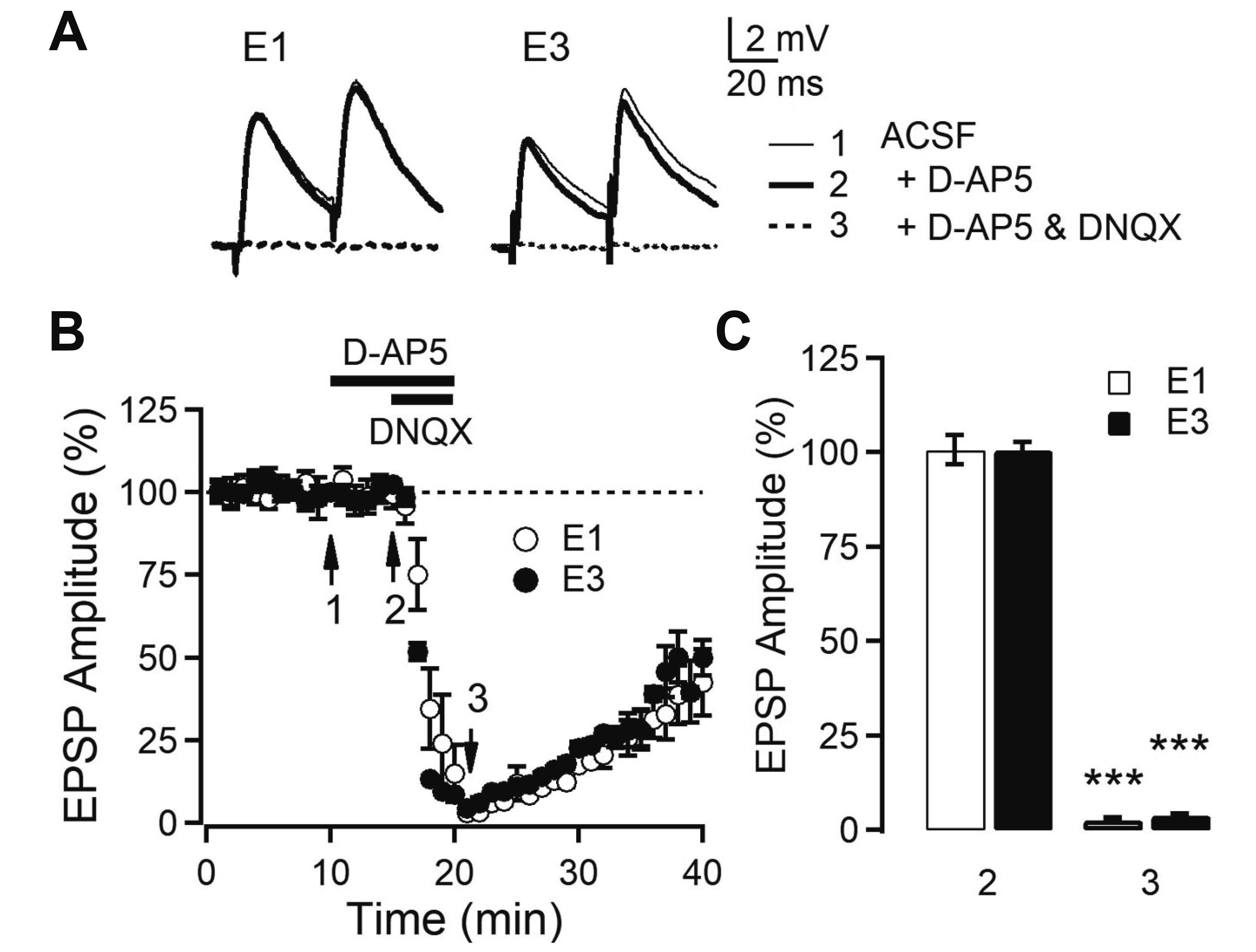 | Fig. 2AMPA receptor-mediated synaptic transmission in Layer 2/3 (L2/3) pyramidal neurons of orbitofrontal cortex.(A) Representative EPSPs evoked by L1 and L3 stimulation (E1 and E3, respectively). The average EPSP waveforms recorded during 1 min at the time indicated by numbered arrows in B. (B) Time course of EPSP amplitude with the application of blockers. The amplitude of first EPSPs evoked by paired-pulse stimulation in the presence and absence of D-AP5 (50 μM) and/or DNQX (20 μM) in bath solution. (C) Summary plot of EPSP amplitude by E1 (n = 6) and E3 (n = 5). ***p < 0.001 compared with baseline. EPSP, excitatory postsynaptic potential; D-AP5, d-(-)-2-amino-5-phosphonopentanoic acid; DNQX, 6,7-dinitroquinoxaline-2,3-dione disodium salt; ACSF, artificial cerebrospinal fluid.
|
Layer-specific serotonergic modulation of glutamatergic transmission in L2/3 PyNs
AMPA receptor-mediated EPSPs were recorded by applying paired-pulse electrical stimulation (40-ms interval) of L1 and L3 alternating in a single PyN. During bath application of 5-HT (10 µM, 10 min) the amplitude of L1-EPSPs decreased by ~30% and then recovered to the baseline level 30–40 min after washout (98.6% ± 1.8%, n = 9, p = 0.632) (Fig. 3A, B). Conversely, the amplitude of L3-EPSP decreased from the baseline to 83.0% ± 2.9% (n = 9, p < 0.001). Therefore, serotonergic stimulation induced input-specific LTD activated by L3 stimulation, but not by stimulation. PPR was not changed in L1-EPSP (1.37 ± 0.06 to 1.33 ± 0.06, n = 9, p = 0.163) and L3-EPSP (1.29 ± 0.11 to 1.32 ± 0.10, n = 9, p = 0.561) (Fig. 3B). In a previous study, combining 5-HT application with tetanic stimulation induced ~25% LTD in the PFC [25]. In the present study, we also applied tetanic stimulation at the end of 5-HT treatment, which induced a rather smaller magnitude of LTD (91.8% ± 1.6% from the baseline, n = 6, p < 0.01). To further investigate the activity dependence of LTD, the test stimulation was turned off during the application of 5-HT and for 10 min after washout (Fig. 3A, B) [7,31]. Without synaptic activity during 5-HT application, no LTD was induced in L3-EPSP (99.1% ± 2.4%, n = 6, p = 0.736) or L1-EPSP (99.5% ± 4.86%, n = 6, p = 0.931). These results indicate that 5-HT induced LTD in a layer-specific and an activity-dependent manner. The concentration-dependent response of 5-HT to L3-EPSP yielded EC50 values of 3.57 µM (n = 5–9) (Fig. 3C). Thus, 10 µM concentration of 5-HT was used to investigate the cellular mechanism of L3 LTD in subsequent experiments.
 | Fig. 3Serotonergic induction of long-term depression (LTD) in the Layer 2/3 (L2/3) pyramidal neurons of orbitofrontal cortex.(A) Time course of average EPSP amplitude evoked by L1 and L3 stimulation. Serotonin (5-HT; 10 μM, 10 min) was applied in bath solution (closed symbols: n = 9). In some experiments electrical stimulation was omitted during the application of 5-HT and until 10 min after washout (open symbols: n = 6). Insets show average EPSPs taken at the time indicated from a representative experiment. (B) Left panels show individual data and averages (thick solid lines). Right panels show the change in paired-pulse ratio (PPR) of closed symbols of the left panels. **p < 0.01 compared with the baseline and ##p < 0.01. (C) Concentration response plot of EPSP amplitude with L3 stimulation. The curve was fitted by sigmoid function (EC50 = 3.57 μM). ***p < 0.001 compared with the normal ACSF. EPSP, excitatory postsynaptic potential; ACSF, artificial cerebrospinal fluid.
|
5-HT2 receptor mediates serotonergic LTD induction
The 5-HT1A and 5-HT2A receptors are the most prevalent 5-HT receptor subtypes in PyNs of the PFC [10]. In the following experiment, we investigated the 5-HT receptor subtypes involved in layer-specific serotonergic induction of LTD using specific antagonists and agonists for 5-HT1A, 5-HT2 and 5-HT3 receptors (Fig. 4). The 5-HT1A receptor agonist 8-OH-DPAT (10 µM) and the 5-HT3 receptor agonist mCPBG (10 µM) failed to induce LTD in L3-EPSP (100.1% ± 2.5%, n = 6, p = 0.969; 99.0% ± 2.3%, n = 6, p = 0.730; respectively). In contrast, the 5-HT2 receptor agonist α-me-5-HT (10 µM) mimicked the LTD induction by 5-HT (82.1% ± 3.4%, n = 6, p < 0.01). Both NAN-190 (10 µM), a 5-HT1A receptor antagonist (87.9% ± 3.6%, n = 5, p < 0.05) and ondansetron (10 µM), a 5-HT3 receptor antagonist (84.9% ± 4.6%, n = 6, p < 0.05), failed to inhibit LTD induced by 5-HT application, respectively. However, 5-HT induction of L3-LTD is blocked by ketanserin (10 µM), a 5-HT2 receptor antagonist (98.1% ± 2.5%, n = 7, p = 0.474). Moreover, ketanserin abolished the effect of α-me-5-HT (98.1% ± 2.3%, n = 6, p = 0.444). Neither 5-HT receptor agonists nor antagonists changed the amplitude of L1-EPSP (Fig. 4A). These results suggest that 5-HT2 receptors mediate the serotonergic induction of L3-LTD in L2/3 PyNs of the OFC.
 | Fig. 45-HT2 receptor mediates the effect of 5-HT induction of long-term depression (LTD).Summary plots of average EPSP amplitude with L1 (A) and L3 (B) stimulation of Layer 2/3 (L2/3) pyramidal neurons of orbitofrontal cortex. 5-HT antagonists were applied into the bath throughout the experiments. 5-HT (n = 9), α-me-5-HT (n = 6), 8-OH-DPAT (n = 6), mCPBG (n = 7), α-me-5-HT + ketanserin (n = 6), ketanserin (n = 7), NAN-190 (n = 5), and ondansetron (n = 6). 5-HT, serotonin; 8-OH-DPAT, 8-hydroxy-2-dipropylaminotetralin hydrobromide; mCPBG, 1-(m-chlorophenyl) biguanide hydrochloride; α-me-5-HT, α-methyl-5-hydroxytryptamine; EPSP, excitatory postsynaptic potential. *p < 0.05, **p < 0.01, ***p < 0.001 compared with the baseline.
|
LTD induction is dependent on intracellular Ca2+ via NMDA receptors
We next investigated the involvement of intracellular Ca2+ and NMDA receptors in LTD induction. Intracellular application of BAPTA (10 mM), a Ca2+ chelating agent, inhibited L3-LTD induced by 5-HT (99.2% ± 1.8%, n = 6, p = 0.681) while L1-EPSP was not changed (100.6% ± 3.0%, n = 6, p = 0.830) (Fig. 5A). NMDA receptor antagonist D-AP5 (50 µM) inhibited L3-LTD (98.6% ± 2.1%, n = 6, p = 0.547), but not L1-EPSP (98.2% ± 1.0%, n = 5, p = 0.157) (Fig. 5B). These results suggest that the increase in intracellular Ca2+ via NMDA receptors might mediate 5-HT induction of L3-LTD. We also determined whether endocannabinoids (eCBs) were involved in the serotonergic induction of LTD in the OFC, since eCBs generated by the increase in intracellular Ca2+ mediated muscarinic LTD in perisomatic synapses of the visual cortex [8]. Application of the cannabinoid (CB) receptor antagonist AM251 (5 µM) failed to inhibit L3-LTD (83.7% ± 4.5%, n = 6, p < 0.05), while the amplitude of L1-EPSP was not changed (97.5% ± 3.4%, n = 5, p = 0.517) (Fig. 5C). These data suggest that eCB signaling played no role in serotonergic LTD in L2/3 PyNs of the OFC.
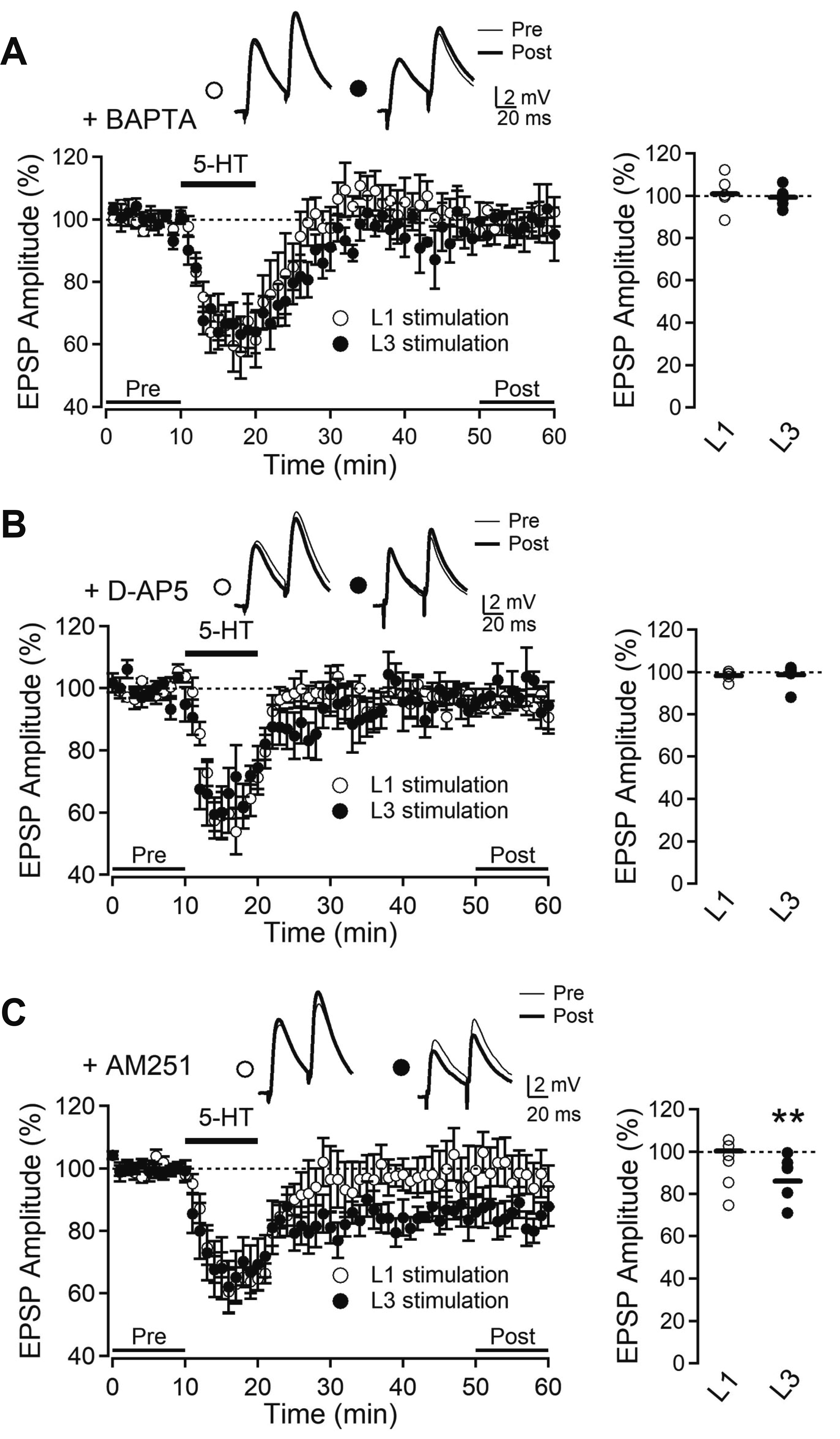 | Fig. 5Serotonergic long-term depression (LTD) is dependent on intracellular Ca2+ and NMDA receptors.(A) Time course of average EPSP amplitude in the presence of intracellular BAPTA (10 mM) with L1 stimulation (open circle, n = 6) and L3 stimulation (closed circle, n = 6). (B) Time course of average EPSP amplitude with bath application of D-AP5 (50 μM) evoked by L1 stimulation (open circle, n = 5) and L3 stimulation (closed circle, n = 6). (C) Time course of average EPSP amplitude with bath application of AM251 (5 μM) evoked by L1 stimulation (open circle, n = 5) and L3 stimulation (closed circle, n = 6). Insets show average EPSPs taken at the time indicated from a representative experiment. Right panels show EPSP amplitude in individual experiments and the average (solid line). EPSP, excitatory postsynaptic potential; BAPTA, 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid; D-AP5, d-(-)-2-amino-5-phosphonopentanoic acid; AM251, N-(piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide. **p < 0.01 compared with baseline (pre).
|
Cholinergic induction of LTD
CCh (10 µM, 10 min) was bath-applied to induce cholinergic LTD and compared with our previous results in the primary visual cortex [7,8]. CCh decreased the amplitude of the baseline amplitude of L1-EPSP to 81.1% ± 3.4% (n = 5, p < 0.01) and L3-EPSP to 86.8% ± 2.5% (n = 6, p < 0.01), respectively (Fig. 6). PPR in L1-LTD (1.11 ± 0.01 to 1.23 ± 0.06, n = 5, p = 0.091) and in L3-LTD (1.04 ± 0.09 to 1.13 ± 0.10, n = 6, p = 0.210) remained unchanged, indicating postsynaptic form (Fig. 6B). The muscarinic ACh receptor (mAChR) blocker atropine (10 µM) in the bath solution blocked the induction of L1-LTD (97.4% ± 2.8% of the baseline, n = 7, p = 0.389) and L3-LTD (98.7% ± 1.4% of the baseline, n = 5, p = 0.454). Therefore, the activation of ACh receptors induced muscarinic LTD in both synaptic inputs on distal apical and perisomatic dendrites in L2/3 PyN of the OFC, similar to the visual cortex. However, contrary to layer-specific serotonergic induction of LTD, cholinergic stimulation induced LTD in both inputs in the OFC.
 | Fig. 6Cholinergic induction of long-term depression (LTD).(A)Time course of average EPSP amplitude evoked by L1 and L3 stimulation. Carbachol (CCh, 10 μM) was applied in bath solution (closed circle: n = 9, closed triangle: n = 6). Atropine (10 μM) was applied to the bath solution throughout the experiment (open circle: n = 7, open triangle: n = 5). Insets show average EPSPs taken at the time indicated from a representative experiment. (B) Left panels show individual data and averages (thick solid lines). Right panels show the change in paired-pulse ratio (PPR) of closed symbols of the left panels. EPSP, excitatory postsynaptic potential. **p < 0.01 compared with the baseline, #p < 0.05, ##p < 0.01.
|
Go to :
DISCUSSION
We investigated layer-specific serotonergic and cholinergic modulation of synaptic transmission in L2/3 PyNs of the OFC. FM1-43 dye unloading demonstrated that the local electrical stimulation of L1 and L3 mainly activated inputs located in L1 and L2/3, respectively. Postsynaptic L3-LTD was induced by 5-HT but not in L1 inputs. 5-HT2 receptors, NMDA receptors, and increase in intracellular Ca2+ mediated L3-LTD. However, the cholinergic stimulation induced muscarinic LTD in both inputs activated by L1 and L3 stimulation. These results suggest that layer-specific neuromodulation of synaptic transmission might be an important mechanism in integration of information in L2/3 PyNs of the PFC.
Layer-specific activation of synaptic transmission in L2/3 PyNs of the PFC
In L2/3 PyNs of the PFC, distal apical dendrites in L1 receive nonspecific thalamocortical inputs whereas the perisomatic dendritic area including basal dendrites in L2/3 receive various synaptic inputs from other cortex, striatum, amygdala, and hippocampus [17-21]. Thus, L2/3 PyNs are well suited for the integration of these diverse synaptic inputs. In the present study, we used local electrical stimulation at L1 and L3 to activate synapses located in distal apical and perisomatic dendrites, respectively. Although we demonstrated that local electrical stimulation activated nearby dendritic synapse in the primary visual cortex in our previous study [8], it was not clear whether this experimental configuration could be applied in the PFC. In the present study, we demonstrated that local electrical stimulation of L1 and L3 activated synapses located in distal apical and perisomatic dendritic areas, respectively, using FM1-43 dye unloading from axon terminals (Fig. 1A–C). Consistently, rapid dynamics of L3-EPSP than L1-EPSP coincided with the distance of synaptic location of respective dendrites from the soma. Furthermore, we showed that L1-EPSP and L3-EPSP were independently evoked by the corresponding stimulation (Fig. 1D). Thus, we established that local electrical stimulation of L1 and L3 independently activated synapses located in distal apical and perisomatic dendrites, respectively, in L2/3 PyNs of the PFC. Inputs to L2/3 PyNs from other brain areas are segregated and terminated onto different dendritic compartments [1,32]. Various active membrane properties are specific to dendritic locations and compartments [33]. Thus, the local electrical stimulation could be an efficient experimental technique to investigate the integration properties of the PFC as well as the primary visual cortex.
Serotonergic LTD and involvement of 5-HT2 receptors in L2/3 PyNs
In the PFC, 5-HT is involved in various forms of long-term synaptic plasticity. LTP in hippocampo-mPFC was augmented in 5-HT-depleted rats [28]. Combining 5-HT treatment with tetanic stimulation induced LTD in layers 5/6 PyNs of the PFC [25]. A 20-min exposure of 5-HT has been reported to induce LTD via activation of 5-HT2A receptors in L5 PyNs of the mPFC [34]. In our previous study, 5-HT inhibited LTP induced by theta-burst stimulation and LTD induced by low-frequency stimulation during the critical period in L2/3 PyNs of the primary visual cortex [35,36]. However, 5-HT reinstated the induction of NMDA receptor-dependent LTP in 8- to 10-week-old rats [37]. In the cat visual cortex, 5-HT promotes the probability of LTP in 5-HT2C receptor-rich zones and facilitates LTD induction in 5-HT2C receptor-poor zones [38]. Thus, 5-HT induces LTP and LTD in a complex manner, depending on the different 5-HT receptor subtypes, brain regions and developmental stages [24,39,40]. In the present study, we demonstrated that a 10-min application of 5-HT (10–30 µM) induced LTD with a 17% decrease in amplitude in the OFC. In a previous study, 5-HT (40 µM) alone reduced excitatory postsynaptic current (EPSC) amplitude in L5 PyNs of the PFC less than 10%, whereas 5-HT treatment combed with tetanic stimulation decreased EPSC amplitude by 25% [25]. In the present study, we also applied tetanic stimulation combined with 5-HT (10 µM), although a smaller amplitude LTD (91.8%) was obtained. Since 5-HT expression varies in receptor subtypes and density depending on cortical areas and layers [40-42], the difference might be attributed to the different areas (mPFC vs. OFC) and layers (L2/3 vs. L5 PyNs).
Among the 14 different 5-HT receptor subtypes, 5-HT1A, 5-HT2A and 5-HT3 are the most abundantly expressed receptors in the PFC. 5-HT2 receptors are coupled to phospholipase C (PLC) and increase intracellular Ca2+. The 5-HT2A/C receptors are highly expressed at the proximal dendrites including the soma according to mRNA expression and immunohistochemical localization [40,42]. In previous studies, 5-HT2 receptor-induced LTD in L5 PyNs of the mPFC was dependent on metabotropic glutamate receptors, in synapses activated by stimulation of either L5 [25] or L1 [34]. Spike timing-dependent LTD (t-LTD) in L1 synapses of L5 PyNs of the mPFC was induced by the activation of presynaptic NMDA receptors upon the activation of presynaptic 5-HT2A receptors [43]. In the present study, 5-HT-induced LTD was mediated by 5-HT2 receptor activation in L2/3 PyNs of the OFC. The 5-HT-induced LTD was NMDA receptor-dependent. We demonstrated that NMDA receptors did not contribute to the peak EPSP amplitude of 5–8 mV upon stimulation of L1 and L3. However, L3-EPSPs showed faster decay kinetics in the presence of D-AP5 in some PyNs (Fig. 2). Since the activation kinetics of NMDA receptors is slower than that of AMPA receptors [44], the small number of NMDA receptors might be activated in perisomatic dendrites by L3 stimulation. Furthermore, the local EPSP amplitude of distal synapses might be large enough to activate NMDA receptors especially in spine heads [45,46]. Moreover, silencing synaptic activation during 5-HT application failed to induce LTD. Thus, the co-activation of 5-HT2 receptors and NMDA receptors is critical for the induction of LTD in OFC L2/3 PyNs. The discrepancy between previous studies in the mPFC and the present study might be attributed to the different cortical areas and layers [47].
In the current study, chelating intracellular Ca2+ with BAPTA blocked L3-LTD. Influx of extracellular Ca2+ from NMDA receptors under 5-HT2 receptor activation could be the source of increased intracellular Ca2+. Otherwise, PLC activation by 5-HT2 receptor-coupled Gq protein could increase intracellular Ca2+ via release from IP3-sensitive stores. In the primary visual cortex, 5-HT2 receptor activation triggered Ca2+ release in the soma and the IP3-mediated facilitation of inhibitory currents was restricted to the perisomatic area, but not L1 [9]. Thus, this previous findings suggest that Ca2+ release from IP3-sensitive stores via activation of 5-HT2 receptors might lead to the induction of L3-LTD in the OFC.
Cholinergic LTD in the prefrontal cortex
Cholinergic regulation of synaptic plasticity in the brain is associated with essential brain functions such as information processing, learning, memory, and attention [31,48,49]. LTD at glutamatergic synapses of L5 PyNs was mediated by mAChRs in the adult mPFC [26,27]. In the present study, in contrast to layer-specific 5-HT induction of LTD, muscarinic stimulation induced LTD in both inputs activated by L1 and L3 stimulation in L2/3 PyNs of the OFC. The magnitude of LTD (~19%) was similar to that of L3-LTD induced by 5-HT application. In our previous study, the magnitude of muscarinic-induced LTD in perisomatic inputs of the primary visual cortex was ~25% [8]. Thus, these findings suggest that the low magnitude of LTD might be an intrinsic property of L2/3 PyNs in the OFC. Although muscarinic and serotonergic stimulation induced LTD, they showed different layer specificity. Thus, these results suggest that different neuromodulators play a role in the induction of long-term synaptic plasticity depending on their own layer-specific patterns. Differential expression of receptors, ion channels and intracellular signaling molecules between dendritic compartments could contribute to the variable effect of neuromodulators.
Implications of layer-specific neuromodulation of synaptic transmission
Cortical PyNs extend their dendrites to the other layers and integrate the inputs from other brain structures in different layers. The L2/3 and L5 PyNs carry multiple perisomatic dendrites including basal dendrites around the soma and a single apical dendrite extending up to L1 as dendritic tufts [2,33]. These dendrites integrate synaptic inputs as compartments with characteristic membrane distribution of ion channels and intracellular Ca2+ release [7,33]. In L2/3 PyNs of the primary sensory cortex, top-down associative inputs from other cortical areas and bottom-up sensory inputs from the thalamus terminate mainly in distal apical dendrites and basal dendrites, respectively [32]. Likewise, in the PFC, feedback connections from other cortical areas as well as the nonspecific thalamus target dendrites in L1 while inputs from amygdala and hippocampus and feedforward cortical inputs terminate in perisomatic dendrites [18,20,21]. In the mPFC, L1 even receives thalamocortical projection and corticocortical inputs in a sublaminar fashion [19]. Thus, the layer-specific nature of synaptic transmission and plasticity of cortical PyNs is important in understanding the cortical integration of information. It is suggested that neuromodulators control synaptic transmission in a pathway-specific manner depending on brain states such as attention, sleep and mood [50]. We previously demonstrated layer-specific muscarinic and serotonergic modulation of synaptic transmission in the primary visual cortex [7-9]. In the present study, 5-HT induced LTD in perisomatic inputs but not in distal apical inputs located in L1 of the OFC while muscarinic stimulation induced LTD in both inputs. This layer-specific neuromodulation may represent an important mechanism of neuromodulatory control of information processing depending on the brain state.
Go to :
 XML Download
XML Download