

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Cardiovascular diseases (CVD) are the leading cause of death worldwide [1]. Vascular endothelial dysfunction is a crucial factor in the occurrence of CVD. And oxidative stress is considered to be endothelial dysfunction major inducer. In particular, oxidative stress caused by ROS is a critical factor in the pathogenesis of vascular diseases. Thus, identifying more effective antioxidants is a promising strategy to prevent endothelial cell injury, which is of great significance for the prevention, clinical diagnosis, and monitoring of endothelial dysfunction and adverse events of CVD [2].
Antioxidant agents that attenuate the oxidative stress may have therapeutic applications in reducing endothelial cell damage. Many plant constituents exhibit antioxidant activity. For sample, Wu et al. [3] showed that the Polygonum orientale flower extract can protect human umbilical vein endothelial cells (HUVECs) from hydrogen peroxide (H2O2)-triggered oxidative damage by enhancing the activity of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase. 2,3,5,4’-Tetrahydroxystilbene-2-O-β-D-Glucoside (TSG), the characteristic water-soluble component of Polygonum multiflorum Thunb, has shown various pharmacologic activities, including antioxidant, anti-inflammatory, anti-aging, and anti-atherosclerotic effects [4-6]. Neuroprotective effects of TSG against glutamate or H2O2-induced oxidative toxicity were confirmed between cell lines or rodent models [7,8]. Li et al. [9] confirmed that TSG decreased pulmonary aortic endothelial cell inflammatory injury induce by septic-serum via the ROS/MAPK/NF-κB signaling pathway. However, the protective effect of TSG on H2O2-induced oxidative damage remains to be fully elucidated.
H2O2 acts as a signal molecule and second messenger involved in most of the redox metabolism reactions and processes of the cells, and is widely used to establish oxidative stress model [10]. Thus, stimulation with H2O2 is a good strategy for investigating vascular endothelial damage. In this study, we aimed to clarify the TSG effects on gene expression (mRNA and microRNA) related to oxidative stress induced by H2O2 in HUVECs.
Go to : 
METHODS
Materials
HUVECs cell line was purchased from Cell Bank of Type Culture Collection of Chinese Academy of Sciences (Shanghai, China). 2,3,5,4'-tetrahydroxystilbene-2-O-β-D-glucoside (TSG, CAS Number: 82373-94-2, purity: ≥ 98%) and TSG was dissolved in sterile water.
Cell cultures and cell treatment
The cells were cultured in RPMI 1640 with 10% fetal bovine serum. The medium was changed every 2 days and the cells were passaged with trypsin-EDTA. HUVECs were randomly divided into 4 groups: control group, H2O2 group, TSG (20 μg/ml) group and TSG (40 μg/ml) group. Cells in the control group were incubated with normal growth conditions. Those in the H2O2 group were incubated with H2O2 (200 μM) for 2 h and then recovered for 24 h. In the TSG (20 μg/ml, 40 μg/ml) groups, the cells were cultured with the medium containing different concentrations of TSG for 24 h before they were treated same as H2O2 group.
MTS assay
The MTS assay was used to assess cell viability. Before each experiment, HUVECs (5 × 103 cells/well) were seeded in 96-well microtiter plates, and cells were treated with H2O2 or TSG according to the different experimental purposes. Subsequently, 20 μl MTS solution was added to each well, and the plates were incubated for 2 h. The absorbance was measured at 490 nm and used to calculate the relative ratio of cell viability.
In situ fluorescence detection of apoptosis
An annexin V-FITC apoptosis detection kit (MultiSciences; LiankeBio, Hangzhou, China) was used for the dual staining assay. Briefly, cells were harvested and washed with PBS. After adding the staining solution per the requirements of the kit, incubate with aluminum foil paper at room temperature for 15 min in the dark, and then place them in an ice bath in the dark. Then observed under a fluorescence microscope within one hour, Annexin V-FITC showed green fluorescence and propidium iodide (PI) showed red fluorescence.
Detection of SOD
We used Total Superoxide Dismutase Assay Kit with WST-8 (Beyotime Biotechnology, Shanghai, China) to detect the SOD activity in cell lysates. After treatments, cell lysates were prepared, and the protein concentration was measured using the BCA assay (Beyotime Biotechnology). The SOD activity levels were quantified according to the manufacturer’s instructions.
RNA isolation and quantitative PCR (qRT-PCR)
Total RNA was extracted with TRIzol reagent. The complementary DNA was synthesized using Prime Script TM RT Master Mix (Perfect Real Time) (Takara, Dalian, China) for real-time polymerase chain reaction (RT-PCR) with conditions of 37°C for 15 min, 85°C for 5 sec, and storage at 4°C. The RT-PCR was performed in duplicate using SYBR®Prime Ex Taq TM II (Tli RNaseH Plus) (Takara) at 95°C for 10 sec, and then 95°C for 5 sec, 60°C for 20 sec for 40 cycles, and 95°C for 1 min, 55°C for 30 sec, 95°C for 30 sec. The primer sequences used in this study were as Table 1 showed.
Table 1
The primer sequences
![]()
TargetScanHuman database
TargetScanHuman database is a miRNA target gene prediction website for human. We inputed the effective miRNA based on the results of qRT-PCR to predict possible target genes.
Statistical analysis
GraphPad Prism 5 software was used for statistical analysis. Measurement data were compared using a one-way analysis of variance. p < 0.05 was considered statistically significant.
Go to :
RESULTS
H2O2 exposure induced the decrease of cell viability
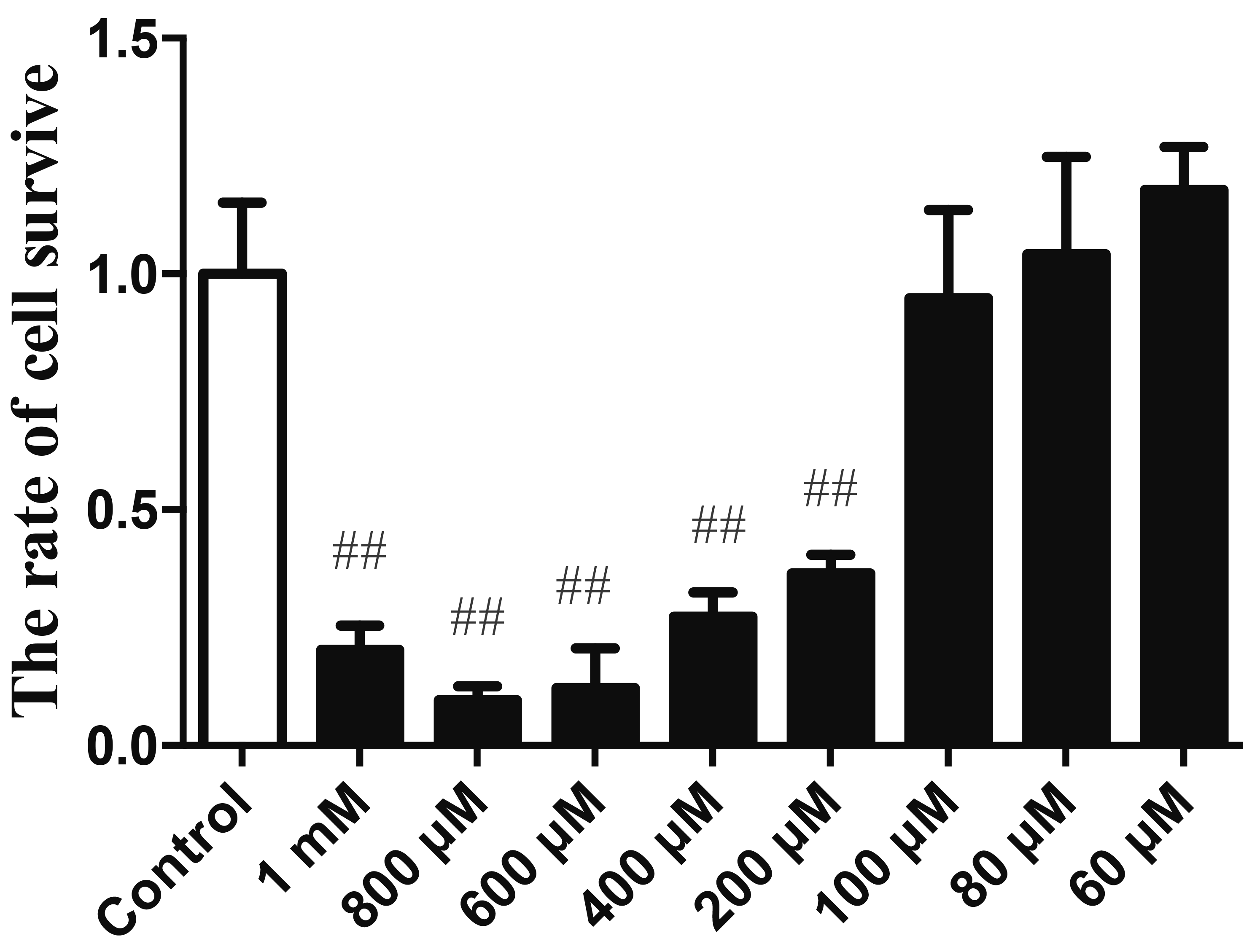
To examine the cytotoxicity of H2O2 on HUVECs, cell viability was detected by the MTS assay. As shown in Fig. 1, treatment with H2O2 (60 μM, 80 μM, and 100 μM) for 12 h had no effect on cell viability of HUVECs. However, cell viability of HUVECs decreased when the concentration of H2O2 increased to 200 μM. Therefore, the concentrations of 200 μM were chosen for H2O2, respectively, for the subsequent experiments.
TSG pre-treatment improved cell viability
TSG treatment at the concentrations of 120 μg/ml, 100 μg/ml, 80 μg/ml, 60 μg/ml, 40 μg/ml, and 20 μg/ml showed no cytotoxicity on HUVECs when compared with the control group (Fig. 2A). Furthermore, the decreased cell viability of HUVECs induced by H2O2 was improved with TSG treatment (40 μg/ml and 20 μg/ml), suggesting that TSG rescued H2O2-induced cell injury in HUVECs (Fig. 2B).
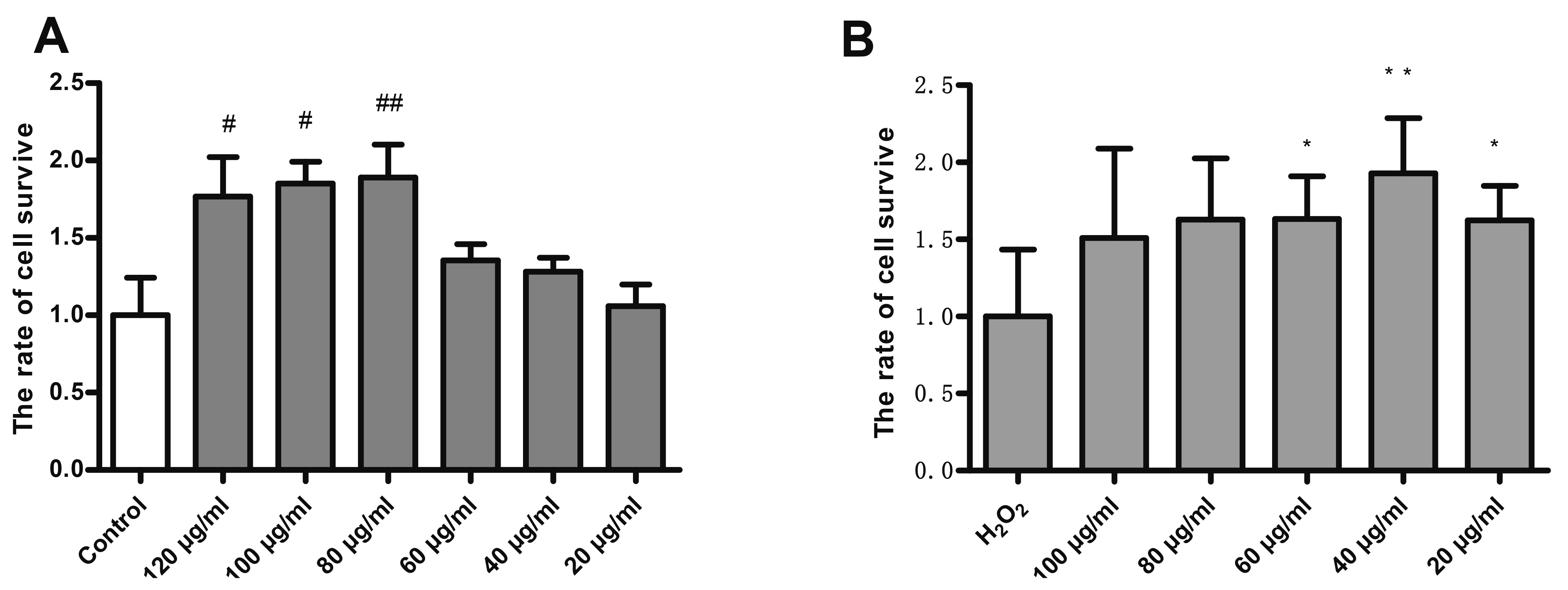 | Fig. 2TSG pre-treatment improved cell viability.TSG Treatment on HUVECs untreated (basal) (A) and H2O2-induced (B) for 24 h. Values are expressed as mean ± standard deviation (n = 3). TSG, 2,3,5,4’-Tetrahydroxystilbene-2-O-β-D-Glucoside; HUVECs, human umbilical vein endothelial cells. #p < 0.05 and ##p < 0.05 vs. control; *p < 0.05 and **p < 0.05 vs. H2O2 group.
|
TSG pre-treatment suppressed apoptotic cell death
Cell death (annexin-V/PI) was explored in HUVECs treated 24 h with TSG in the presence or absence of H2O2 (200 µM). H2O2 can induce various cell death, including apoptosis and necrosis. We measured the Annexin V positive and PI positive HUVECs of each group to determine the state of the cells. As shown in Fig. 3, the treatment of HUVECs with 200 μM H2O2 significantly increased the apoptotic cell death. And pre-incubation with TSG significantly attenuated H2O2-induced apoptotic cell death.
 | Fig. 3TSG pre-treatment suppressed apoptotic cell death.Representative images of staining with Annexin V-FITC (green) and PI (red) acquired using a laser scanning microscope (scale ×100 μm). The images were captured within the same field and then merged. Values are expressed as mean ± standard deviation (n = 3). TSG, 2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucoside; PI, propidium iodide.
|
TSG pre-treatment ameliorated H2O2 induced loss of SOD in HUVECs
SOD is an important metal enzyme that can specifically remove superoxide anions and protect the human body from oxidative damage. Moreover, SOD is a key mitochondrial antioxidant. We measured the intracellular SOD activity of each group to determine the extent of their oxidative damage. The result of the antioxidant system of SOD was shown in Fig. 4. The activity of SOD significantly decreased in H2O2-treated group compared with that in the control group, and was significantly actived by TSG pre-treatment with increasing concentrations.
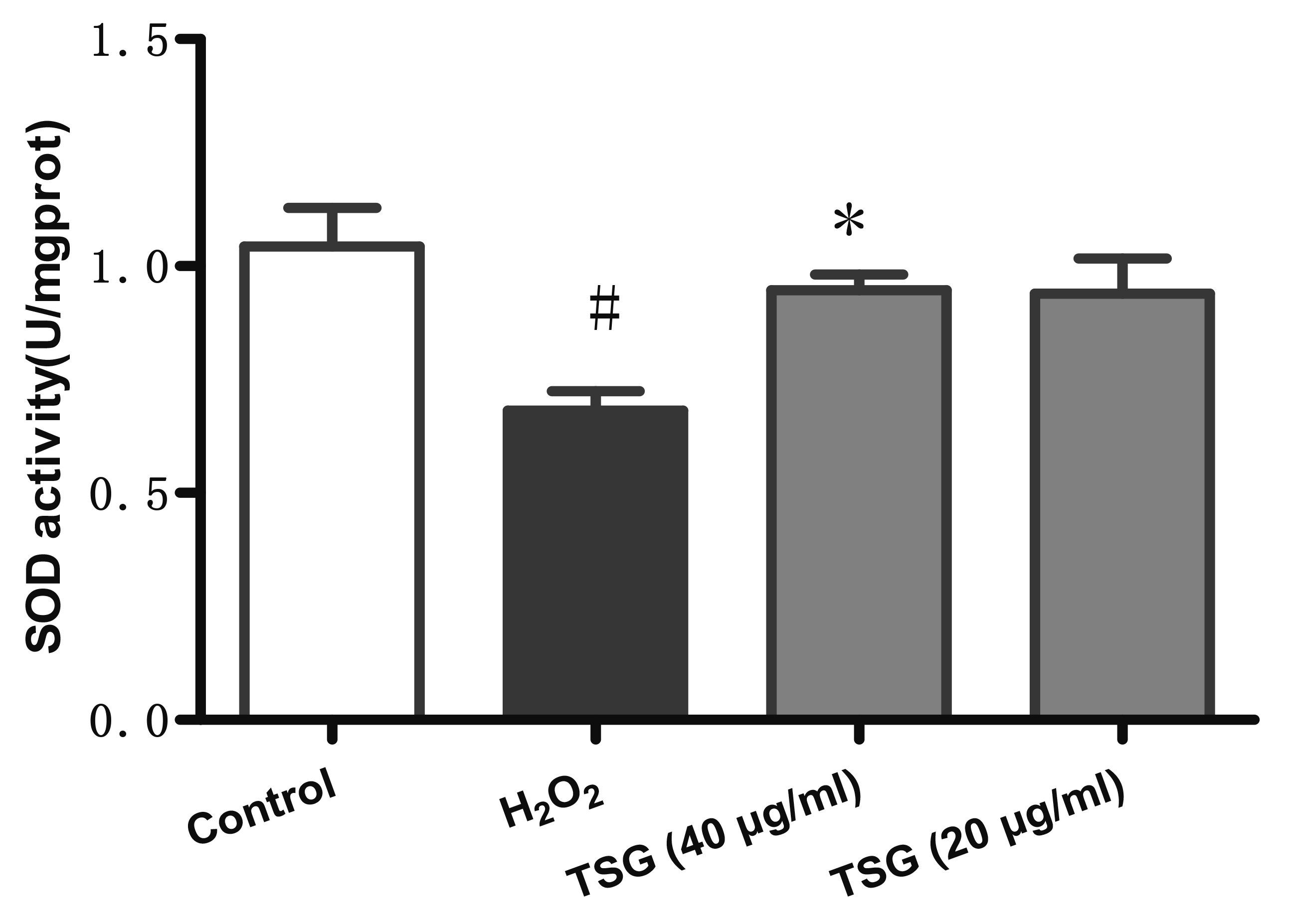 | Fig. 4TSG pre-treatment ameliorated H2O2 induced loss of SOD in HUVECs.SOD activity was been detected. Values are expressed as mean ± standard deviation (n = 3). TSG, 2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucoside; SOD, superoxide dismutase; HUVECs, human umbilical vein endothelial cells. #p < 0.05 vs. control; *p < 0.05 vs. H2O2 group.
|
Effect of TSG pre-treatment in miRNA expression in H2O2 induced HUVECs
Studies have shown that miRNA can regulate the expression of most genes in cells. In the H2O2-treated HUVECs, the expression of miR-9-5p, miR-16, miR-21, miR-29b, miR-145-5p, and miR-204-5p were significantly upregulated. TSG antagonized H2O2-induced miR-9-5p, miR-16, miR-21, miR-29b, miR-145-5p, and miR-204-5p synthesis (Fig. 5A–F).
 | Fig. 5Effect of TSG pre-treatment in miRNA expression in H2O2 induced HUVECs.qRT-PCR was used to detected the (A) miR-9-5p, (B) miR-16, (C) miR-21, (D) miR-29b, (E) miR-145-5p, and (F) miR-204-5p synthesis. Values are expressed as mean ± standard deviation (n = 3). TSG, 2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucoside; HUVECs, human umbilical vein endothelial cells. #p < 0.05 and ##p < 0.01 vs. control; *p < 0.05 and **p < 0.01 vs. H2O2 group.
|
Effect of TSG on endoplasmic reticulum (ER) stress in HUVECs
The ER has recently emerged as an alternative target to induce cell death, because prolonged ER stress results in the induction of apoptosis. The levels of ER stress markers (ATF4, ATF6, CHOP, GRP78) and E2F1 mRNA were examined with qRT-PCR. In the H2O2-treated HUVECs, the expression of ATF6 and CHOP mRNA was significantly upregulated while the ATF4, GRP78 and E2F1 mRNA were significantly downregulated. TSG antagonized H2O2-induced ATF6, CHOP and E2F1 mRNA synthesis. However, TSG has no effects on reversing the expression of ATF4 and GRP78 mRNA (Fig. 6A–E).
 | Fig. 6Effect of TSG on ER stress in HUVECs.qRT-PCR was used to detect the (A) ATF4, (B) ATF6, (C) CHOP, (D) GRP78, and (E) E2F1 mRNA synthesis. Values are expressed as mean ± standard deviation (n = 3). TSG, 2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucoside; ER, endoplasmic reticulum; HUVECs, human umbilical vein endothelial cells. #p < 0.05 and ##p < 0.01 vs. control; *p < 0.05 and **p < 0.01 vs. H2O2 group.
|
Go to :
DISCUSSION
CVD are major contributor to death associated with endothelial dysfunction. Effective drugs to prevent endothelial dysfunction are important. In the present research, our results showed that TSG could mitigate H2O2-induced decrease of cell vitalities and antioxidative enzyme activities and decrease the level of oxidative stress and ER stress in HUVECs. And the safety of TSG has been well proved. Our result showed that TSG at the concentration had no effect on the cell viabilities of HUVECs. Notably, the concentration of 40 μg/ml and 20 μg/ml showed a property to inhibit H2O2-induced oxidative stress and ER stress.
SOD is an important metal enzyme that can specifically remove superoxide anions and protect the human body from oxidative damage. It is closely related to the occurrence and development of many diseases. After we measured the total SOD activity in cells, we found that TSG can reduce the oxidative damage of H2O2-induced HUVECs by increasing the SOD activity in cells. In addition, the results of in situ fluorescence detection of apoptosis also visually show that TSG significantly inhibits H2O2-induced HUVECs apoptotic cell death.
miRNAs are small endogenous non-coding RNAs of 18–24 nucleotides in length, which are widely present in plants and animals and are involved in various biochemical regulations of the body, including cell proliferation, senescence, death, cell cycle regulation and apoptosis [11-13]. For example, studies have investigated the role of miR-9-5p in angiogenesis and apoptosis in HUVECs injury, and dual luciferase reporter gene assay verified that miR-9-5p targeted CXCR4 [14], while miR-16, miR-29b exerts a specific tumor suppressive effect by targeting the oncogene Bcl-2 [15,16]. As a key regulator of apoptosis in many types of human tumors, Bcl-2 is also positively regulated by miR-21 [17]. In addition, miR-145 has a negative regulatory effect on caspase-3 and inhibits the apoptosis process of cells [18]. Moreover, studies have shown that HUVECs were suppressed by conditioned media from lung cancer cells with miR-204 overexpression [19]. Based on the above research, we explored whether TSG has effects on the expressions of miRNAs in H2O2-induced HUVECs. And the results showed that TSG can play a protective role against HUVECs oxidative damage via reducing the expression of miR-9-5p, miR-16, miR-21, miR-29b, miR-145-5p and miR-204-5p. These may also prompt us to further study the antioxidant mechanism of TSG in the future consider miR-9-5p, miR-16, miR-21, miR-29b, miR-145-5p, and miR-204-5p.
ER stress occurs after oxidative stress. ER represents a compartment able to sense several cellular stresses and to trigger, as last resort, cell death [20]. ER stress markers included the ATF4, ATF6, CHOP, and GRP78. ATF4/ATF6 [21,22] which activates transcription factor (ATF/CREB) family, molecular chaperone GRP78 in the endoplasmic reticulum, and CHOP, a transcription factor that mediates endoplasmic reticulum stress-induced apoptosis [23], P53 [5], etc. have an important relationship with oxidative stress and apoptosis. Our results showed that TSG may exert its protective effect against HUVECs oxidative damage via decreasing the expressions of ATF6 and CHOP. TSG had little effects on the expression of ATF4 and GRP78. Meanwhile, we made use of the TargetScanHuman database to find that ATF6 may be the target gene of miR-16 (Fig. 7).

E2F1 is known to exert different effects on cell growth and apoptosis depending on the cell context. ER stress-mediated E2F1 down-regulation may contribute to the life/death cell decision under prolonged ER stress. Our results indicated that the low expression of E2F1 will promote HUVECs oxidative damage, while TSG can play its protective role against HUVECs oxidative damage via increasing the expression of E2F1. Pagliarini et al. [24] demonstrated that ATF6 controls the expression of E2F1 during the UPR program execution through direct binding to an ERSE site within the E2F1 gene promoter. Therefore, we could hypothesis that TSG regulate the expression of E2F1 through ATF6.
In summary, TSG can effectively protect HUVECs from oxidative damage induced by H2O2. Its main regulatory mechanism may be related to miR16/ATF6/E2F1 signaling pathway. And the molecular mechanism of its specific role remains to be further explored.
Go to :
 XML Download
XML Download