

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
The increasing prevalence of vascular-related diseases such as atherosclerosis, cardiovascular events, retinopathy, and nephropathy during diabetes is a serious life-threatening concern all over the world. Indeed, vascular endothelial cells, vascular smooth muscle cells, and connective tissue cells are at risk of damage to hyperglycemia-induced oxidative stress and the subsequent systemic and local inflammation [1,2]. The available evidence has shown that endothelial dysfunction and adverse vascular remodeling accompanying diabetes are correlated with overexpression of inflammatory mediators along with increased nuclear factor- kappa B (NF-κB) activity [3]. Results of convincing studies have demonstrated that the activated NF-κB signaling pathway in chronic diabetic conditions acts in a vicious cycle to upregulate the inflammatory genes and cytokines like interleukin-1 beta (IL-1β), IL-6, tumor necrosis factor-alpha (TNF-α), and intercellular and vascular cell adhesion molecules (ICAM, VCAM), exacerbating the vascular tissue injury [4].
In addition, microRNAs (miRNAs), non-coding RNAs in the cell cytoplasm, have been known to be a regulator of gene expression through interaction with the complementary section of its target mRNAs, and play crucial physiological roles in cell function hemostasis as a suppressor or enhancer in gene expression. However, if the specific miRNAs gene in a particular tissue were dysregulated by inflammatory mediators, they would take part in many pathological situations like diabetes, influencing various cell signaling and inflammatory modulation [5]. In this sense, miRNA-146a is an important biomarker of inflammatory status which it would be considered as a new therapeutic target in the management of the related disease. So, the assessment of its dysregulation during diabetes can be a reliable approach to study the pathophysiological target pathways, especially the NF-κB signaling pathway [5,6]. Results from previous studies have demonstrated that diabetes impaired the expression of miRNA-146a that led to the high-level activity of NF-κB-related inflammatory signaling and its adaptor proteins like interleukin-1 receptor-associated kinase-1 (IRAK-1) and TNF receptor-associated factor-6 (TRAF-6) in diabetic tissues [7,8]. Thus, the preventive and therapeutic strategies targeting these elements of inflammatory reactions during diabetes would be promising in minimizing the diabetes-induced vascular complications.
Troxerutin (trihydroxy-ethyl rutin) (Fig. 1), a natural bioflavonoid, has been reported to have many medicinal properties including anti-inflammatory, antioxidant, anti-apoptotic, antithrombotic and vasoprotective features, with fewer side effects [9,10]. In a study, troxerutin positively affects the hippocampus function by reducing oxidative stress and promoting learning and memory in streptozotocin-induced type I diabetic rats [11]. Also, previous studies revealed anti-apoptotic and anti-inflammatory effects of troxerutin against myocardial ischemia/reperfusion injury through the prevention of the cell-cell interaction in healthy rats [12], and anti-oxidative effect on aortic tissue of diabetic rat trough enhancing tissue antioxidant system activity and declining lipid peroxidation level [13]. Besides, in another study in healthy rats, troxerutin has exerted a myocardial protective effect against ischemia/reperfusion injury as preconditioning via the anti-apoptosis process by regulating miR‐146a downregulation [12].
Thus, due to the encouraging potentials of troxerutin in alleviating the oxidative-inflammatory conditions in cardiovascular medicine, and the significant roles of vascular inflammatory responses and related miRNA in diabetes pathophysiology, in this study, we investigated the preventive effects of troxerutin on the miRNA-146a/NF-κB signaling pathway, and endothelial and vascular cell inflammatory mediators in aortic tissue of healthy and diabetic rats.
Go to : 
METHODS
Animals and materials
The study was carried out with thirty-two male Wistar rats (7–8 months) weighing 200–230 grams. Animals were kept in polypropylene cages and experienced a 12-h light/12-h dark, 50% humidity and temperature of 25°C and had free access to food and water. This study was approved by the local committee for animal care under ethical number IACUC-2019(1)1025, and the Guidelines for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication No 85-23, revised 1996) were followed throughout the study. Troxerutin was purchased from Sigma (St. Louis, MO, USA) and all other chemicals and reagents were obtained from commercial sources in the highest quality available.
Experimental design and induction of diabetes
Animals were randomly divided into four groups (n = 6 in each group) including healthy control (Healthy-Cont), healthy with troxerutin (Healthy-TXR), diabetic control (Diabetic-Cont), and diabetic with troxerutin (Diabetic-TXR). Type-I Diabetes was induced by a single intraperitoneal injection of streptozotocin (STZ; 60 mg/kg; dissolved in 0.1 molar citrate buffer with pH 4.5). The healthy rats received the same amount of citrate buffer alone. The development of diabetes was confirmed by measuring the blood glucose levels, 72 h after injection. The animals with blood glucose levels higher than 300 mg/dl were considered diabetic and those with blood glucose levels lower than this value were excluded from the experiment [14]. After six weeks of STZ injection, troxerutin (150 mg/kg/day) was administered orally for four weeks [15]. The experimental period lasted ten weeks in all groups.
Sampling and tissue processing
At the end of the experimental duration, the blood glucose in fasted-rats from each group was measured using a glucometer. Then, the animals were weighed and anesthetized with an intraperitoneal injection of 60 mg ketamine and 10 mg xylazine per kg of body weight. The thoracic aorta was immediately removed and rinsed in cold saline and weighed. The aortic tissue was frozen by liquid nitrogen and stored in a –80°C refrigerator. Then the aortic samples were homogenized in lysis buffer at pH 7.4 in the presence of protease inhibitor cocktail (Sigma-Aldrich). Then, the homogenate was centrifuged at 10,000 rpm for 10 min at 4°C. The supernatants were removed from the homogenates and quickly frozen at –80°C. The protein concentration of the supernatant was estimated using the Bradford technique.
Measurement of inflammatory cytokines, enzymes and adhesion molecules
In all aortic tissue, the total RNA was extracted using Alterations of the pro-inflammatory cytokines and mediators in aortic samples of the experimental groups were evaluated using the enzyme-linked immunosorbent assay (ELISA) methods. Rat specific ELISA kits were used to detect the tissue levels of pro-inflammatory cytokines IL-6, IL-1β and TNF-α, and activity of enzymes including inducible nitric oxide synthase (iNOS) and cyclooxygenase-II (COX-II), and adhesion molecules ICAM-1 and VCAM (MyBiosource, Inc., San Diego, CA, USA), according to the kits guidelines provided by the manufacturers. Fifty microliters of each supernatant and standard were mixed with assay buffers in duplicates according to the instructions. The absorbance was measured at 450 nm using a microplate ELISA reader (Lab System, Helsinki, Finland). The optical densities were normalized according to the corresponding standard curves and the resultant values expressed as pg/mg of protein in each sample.
Real-time PCR assessments of mRNAs and miRNA
In all aortic tissue, the total RNA was extracted using the miRCURY RNA Isolation Kit (Exiqon, Vedbaek, Denmark) according to the manufacturer’s instructions. To measurement of the quantity and quality of the isolated RNA were made in a Nano-drop spectrophotometer (Thermofisher Scientific, Waltham, MA, USA). The content of each sample was at least 200 ng/µl. The cDNA synthesis from RNA was performed with the specific cDNA Synthesis kit (Fermentas GmBH, St. Leon-Rot, Germany) with the aid of random hexamer primers and MMLV reverse transcriptase and then the mRNA expressions were obtained using specific primers for each inflammatory mRNAs and miRNA, including NF-κB, IRAK-1 and TRAF-6, miRNA-146a, and housekeeping genes GAPDH and U6. Each cDNA was used as a template for real time PCR analysis employing the SYBR Green master mix (Exiqon, Vedbaek, Denmark), based on the instruction. The forward and reverse primer sets, respectively, for NF-κB were AATTGCCCCGGCAT and TCCCGTAACCGCGTA, for IRAK-1 were GCTGTGGACACCGAT and GCTACACCCATCCACA, for TRAF-6 were CAGTCCCCTGCACATT and GAGGAGGCATCGCAT, for GAPDH were CCCATCACCATCTTCCAGGAG and GAAGGGGCGGAGATGATGAC, and for miRNA-146a was UGAGAACUGAAUUCCAUGGGUU. The PCR reactions were conducted on a Bio-Rad iQ5 Detection System (Bio-Rad, Richmond, CA, USA). The 2–ΔΔCt algorithm was used to determine and analyze the expression levels of each mRNA and miRNA.
Statistical analysis
Data are expressed as means ± standard error. Differences of parameters between the groups were analyzed using one-way ANOVA followed by Tukey’s test, as appropriate. p-value of less than 0.05 was considered statistically significant.
Go to :
RESULTS
Body weights, and plasma levels of glucose and insulin
There was no significant difference in initial body weights among experimental groups (Table 1). After ten weeks of the diabetic period, the final body weight and plasma insulin concentration decreased (p < 0.05) and plasma glucose level increased (p < 0.01) in diabetic control rats in comparison with the healthy control group. Troxerutin in treated-diabetic rats inhibited the diabetes-induced loss of final body weight and considerably reduced the glucose level and significantly increased the plasma insulin level (p < 0.05), as compared with those of diabetic control rats (Table 1).
Table 1
The effect of troxerutin on body weight and blood glucose and insulin levels in healthy and diabetic rats
| Groups | Initial body weight (g) | Final body weight (g) | Blood glucose (mg/dl) | Insulin level (ng/ml) |
|---|---|---|---|---|
| Healthy-Cont | 206 ± 9 | 221 ± 16 | 97 ± 15 | 3.80 ± 1.01 |
| Healthy-TXR | 211 ± 7 | 246 ± 19 | 93 ± 10 | 4.26 ± 1.06 |
| Diabteic-Cont | 214 ± 11 | 183 ± 13a | 426 ± 38b | 1.40 ± 0.30b |
| Diabetic-TXR | 209 ± 8 | 214 ± 17 | 302 ± 27c | 2.36 ± 0.90c |
![]()
Expression of NF-κB, IRAK-1, and TRAF-6 genes in aortic tissue
Gene expression assessment using real-time PCR showed that the mRNA levels of NF-κB, IRAK-1, and TRAF-6 genes in aortic samples of rats were significantly increased in the diabetic group in comparison with the healthy control group (p < 0.05 for NF-κB & IRAK-1; and p < 0.01 for TRAF-6) (Fig. 2). However, pre-administration of troxerutin in diabetic rats significantly reduced the expression of all these genes as compared with the diabetic control group (p < 0.05). In healthy non-diabetic rats, troxerutin significantly decreased only NF-κB expression in comparison to the healthy control rats, but it had no significant effect on IRAK-1, and TRAF-6 genes (Fig. 2). These results demonstrated the suppressing effect of troxerutin on the vascular expression of NF-κB and its adaptor molecules in type-I diabetic rats.
 | Fig. 2Real-time PCR analysis of genes expressions in the aortic tissue.The expression (mRNA) levels of NF-κB (A), IRAK-1 (B) and TRAF-6 (C). The data were expressed as mean ± standard error. n = 6 for each group. NF-κB, nuclear factor kappa B; IRAK-1, interleukin-1 receptor-associated kinase-1; TRAF-6, tumor necrosis factor receptor-associated factor-6; Cont, control; TXR, troxerutin. *p < 0.05, and **p < 0.01 vs. Healthy-Cont group; and #p < 0.05 vs. Diabetic-Cont group.
|
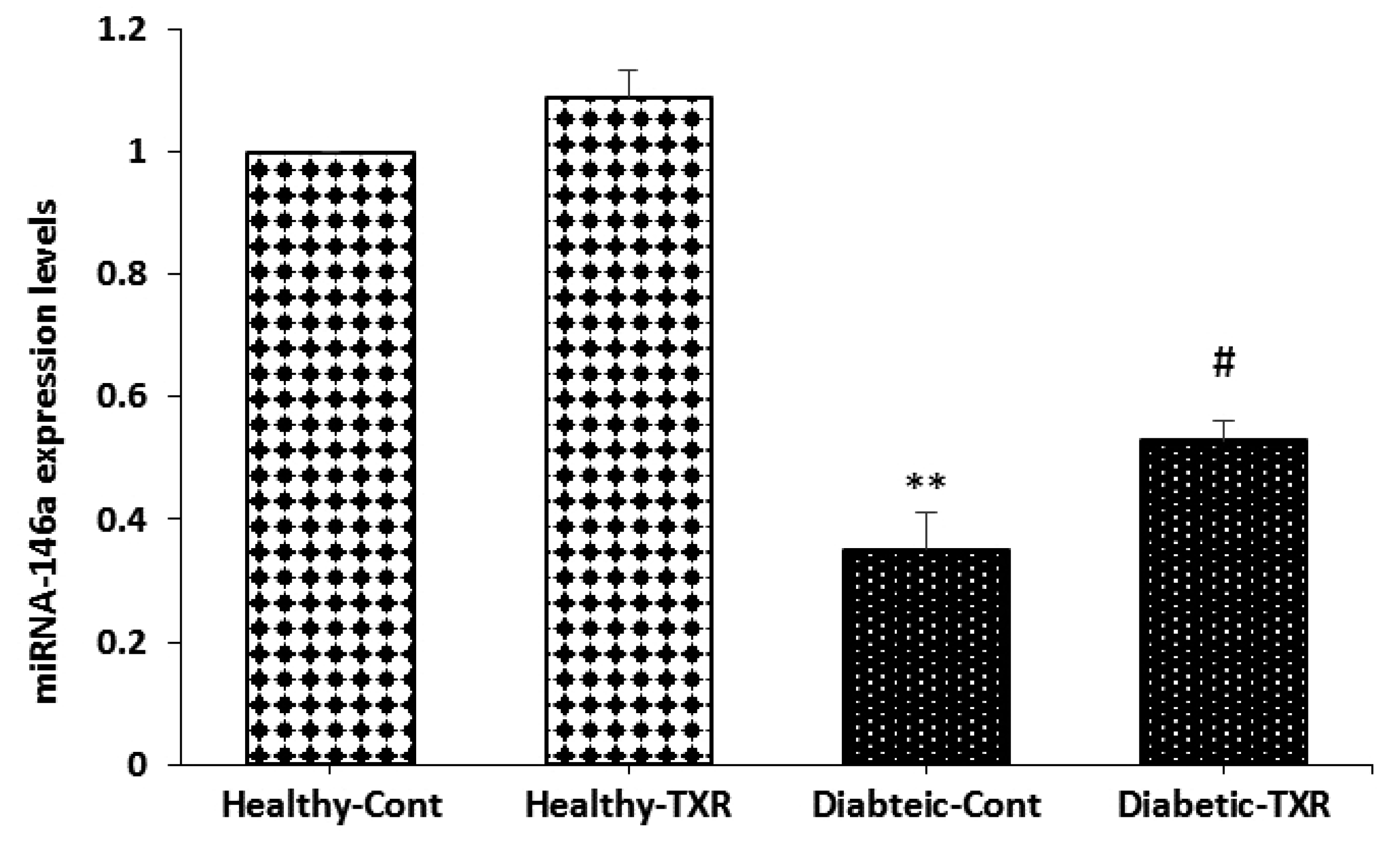
Expression of miRNA-146a in aortic tissue
Induction of diabetes significantly reduced the expression of miRNA-146a as compared with the healthy group (p < 0.05; Fig. 3). The pretreatment of healthy rats with troxerutin for one month could not influence the expression of miRNA-146a when compared to healthy controls. However, in diabetic rats, troxerutin significantly raised the expression of this anti-inflammatory miRNA as compared with the diabetic control group (p < 0.01).
Levels of pro-inflammatory cytokines in aortic tissue
The aortic levels of IL-6, IL-1β, and TNF-α were measured as the main indicators of vascular pro-inflammatory cytokines activity. As shown in Fig. 4, the levels of these cytokines were significantly increased following the induction of diabetes in comparison to the healthy control group (p < 0.05 for IL-6 and p < 0.01 for others). Although administration of troxerutin had significant effects on only TNF-α (p < 0.05) in healthy non-diabetic rats, this bioflavonoid significantly reduced the levels of IL-6, and IL-1β (p < 0.05), and TNF-α (p < 0.01) in treated diabetic group versus diabetic controls (Fig. 4, respectively).
 | Fig. 4The levels of inflammatory cytokines in the endothelial cells of aortic tissue.IL-6 (A), IL-1β (B) and TNF-α (C). The data were expressed as mean ± standard error. n = 6 for each group. IL, interleukin; TNF-α, tumor necrosis factor-α; Cont, control; TXR, troxerutin. *p < 0.05 and **p < 0.01 vs. Healthy-Cont group; and #p < 0.05 vs. Diabetic-Cont group.
|
Levels of adhesion molecules in aortic tissue
ICAM-1, and VCAM are two adhesion molecules which are involved in the interactions between inflammatory and endothelial/vascular cells. Again, induction of diabetes caused the significant elevation of molecules ICAM-1 (p < 0.01) and VCAM-1 (p < 0.05) in comparison to those of healthy control group (Fig. 5, respectively). Pre-administration of troxerutin to the diabetic rats significantly reduced the diabetes-induced elevation of ICAM-1 levels toward healthy control values (p < 0.05).
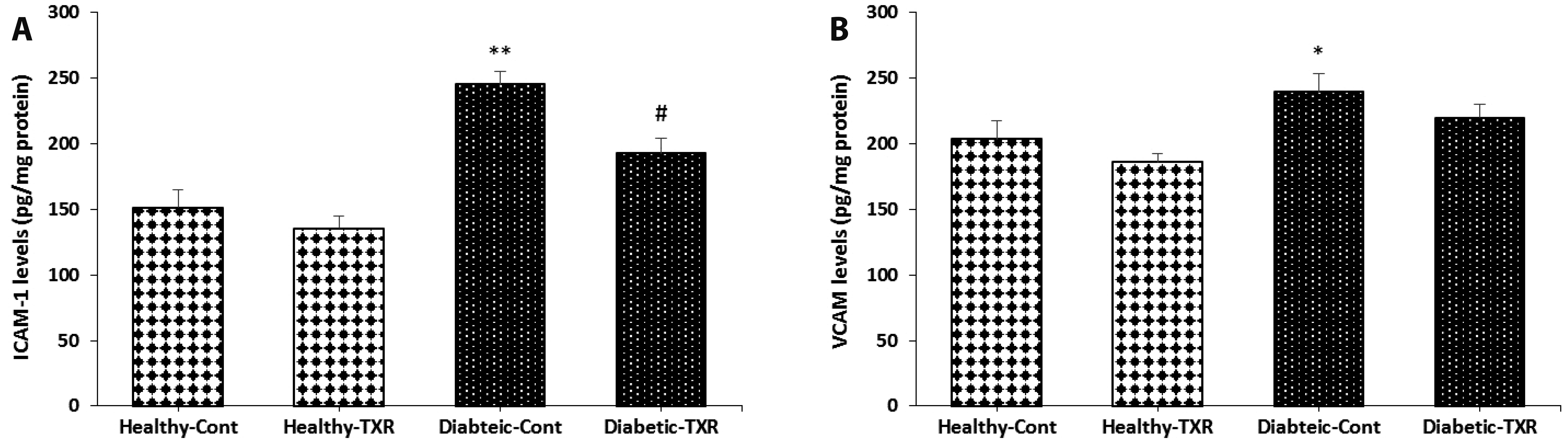 | Fig. 5The levels of adhesion molecules in aortic tissue.ICAM-1 (A), VCAM (B). The data were expressed as mean ± standard error. n = 6 for each group. ICAM-1, intercellular adhesion molecule-1; VCAM, vascular cell adhesion molecule; Cont, control; TXR, troxerutin. *p < 0.05 and **p < 0.01 vs. Healthy-Cont group; and #p < 0.05 vs. Diabetic-Cont group.
|
Levels of iNOS and COX-II in aortic tissue
Besides pro-inflammatory cytokines, two related enzymatic mediators iNOS and COX-II are intracellular targets for the NF-κB transcription factor during inflammatory conditions. In healthy control rats, troxerutin pretreatment significantly reduced the COX-II level (p < 0.05) but not the iNOS level. The levels of these two mediators were significantly elevated by the induction of diabetes as compared with healthy control rats (p < 0.01; Fig. 6). Conversely, pre-treatment of diabetic rats with troxerutin for 4 weeks significantly inhibited the diabetes-induced elevation of both iNOS and COX-II levels as compared with those of the diabetic control group (p < 0.05). Overall, these results confirmed that the anti-inflammatory and endothelial-improving effects of troxerutin on the aortic tissue of diabetic rats are likely achieved by the regulation of the elements of the NF-κB signaling pathway.
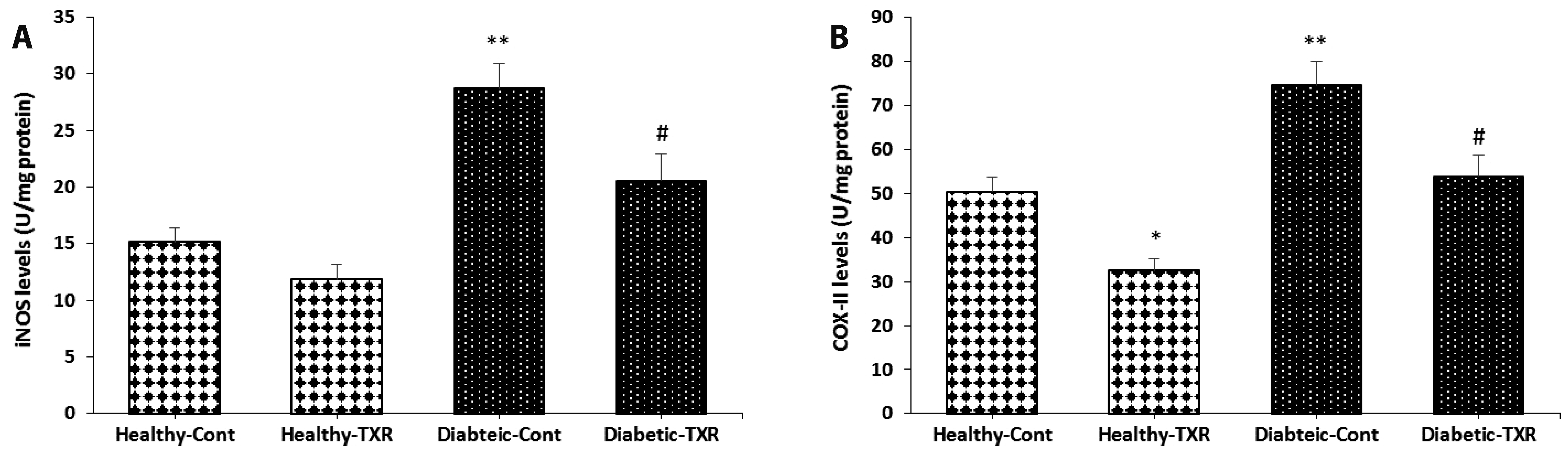 | Fig. 6The levels of inflammatory inducible enzymes in aortic tissue.iNOS (A), and COX-II (B). The data were expressed as mean ± standard error. n = 6 for each group. iNOS, inducible-nitric oxide synthase; COX-II, cyclooxygenase-II; Cont, control; TXR, troxerutin. *p < 0.05 and **p < 0.01 vs. Healthy-Cont group; and #p < 0.05 vs. Diabetic-Cont group.
|
Go to :
DISCUSSION
Findings from this study demonstrated that pretreatment of STZ-induced diabetic rats corrected the hyperglycemia and hypoinsulinemia to some extent and led to a decrease in the expression of IRAK-1, TRAF-6, NF-κB genes that followed by a decrease in pro-inflammatory cytokines (IL-1β, IL-6, TNF-α), inflammatory-related enzymes (iNOS, COX-II), and adhesion molecules (ICAM-1, VCAM), as well as an increase in the expression of miRNA-146a gene in the aortic tissue. Troxerutin administration counteracted these type-I diabetes-induced alterations, indicating its broad and significant anti-inflammatory effects in aortic tissue.
The available data suggest that diabetic-related vascular dysfunctions, especially endothelial impairments, manifest as atherosclerosis, cardiovascular complications, retinopathy, nephropathy and so on [16]. The relevant mechanisms of endothelial dysfunction during diabetes include impaired NO synthase function and reduced NO bioavailability through lack of PI3 kinase/Akt pathway [17,18], increased oxidative stress and inflammatory processes [3,4,16], over-activation of PKCβ [19], and mitochondrial dysfunction [20]. Activated endothelial cells produce inflammatory cytokines such as IL-6, monocyte chemotactic protein-1 and adhesion molecules including ICAM-1, VCAM in arterial tissue [21,22]. It has known that in type-I diabetes, oxidative stress-induced inflammation associates with overexpression of Toll-like receptors on the lymphoid cells of the immune system, which are stimulated by receptor ligands, damage-associated molecular patterns, and inflammatory cytokines, especially TNF-α in damaged cells. The activation of Toll-like receptors activate the NF-κB signaling (IRAK-1/TRAF-6/NF-κB pathway), leading to gene upregulation of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α), intercellular and vascular cell adhesion molecules, inducible enzymes (iNOS and COX-II) that induce more pro-inflammatory state in different cell type and diseases, such as type-I diabetes and atherosclerosis [23,24]. In line with previous studies, STZ-induced diabetes in this experiment caused more inflammatory responses that were established with increasing IL-1β, IL-6, and TNF-α, ICAM-1, VCAM, iNOS, and COX-II accompanying NF-κB pathway activation in the aortic tissue. The administration of troxerutin significantly reversed the diabetes-induced changes. Results of previous studies have confirmed that troxerutin has anti-oxidative and anti-inflammatory functions in different vascular and non-vascular tissue types [9-11]. Although, in the present study, treatment of type-I diabetic rats with troxerutin slightly influenced hyperglycemia, the effects of this bioflavonoid on diabetic-provoked inflammatory conditions were substantial. As previous studies have reported that the anti-inflammatory effect of troxerutin might be accomplished by suppressing NF-κB activation in non-vascular tissues [25], our results revealed the involvement of this mechanism in vascular tissue, so that troxerutin repressed IRAK-1/TRAF-6/NF-κB pathway in aortic and improved the activity of vascular inflammatory mediators.
Recently, the miRNAs-NF-κB pathway has been considered as a new target for manipulating and preventing the concomitant diabetic-induced conditions [26]. The decreased miRNA‐146 expression has been reported in peripheral blood mononuclear cells of patients with type-I diabetes [27,28], and endothelial cells of mice [29]. Similarly, in this study, the improving effects of troxerutin on inflammatory reactions and intercellular adhesion molecules were associated with enhanced expression of miRNA-146a in aortic tissue of type-I diabetic rats. MiRNA-146a has been reported as a potent anti-inflammatory regulator in various diseases associated with inflammation and oxidative stress. Sustained inflammation and oxidative stress in various cell types stimulate the expression of miRNA-146a through activated transcription factor NF-κB. Increased miRNA-146a expression, in turn, could block the expression of its target genes such as IRAK1 and TRAF6, downregulating the activation of Toll-like receptor/NF-κB-mediated inflammation and over-activation of cytokines, iNOS, COX-II and ICAM-1. During the inflammation, the TRAF6/NF-κB pathway is strongly regulated by a negative feedback regulatory loop with miRNA-146a as a key component, serving to protect tissues against inflammatory reactions. Here, the NF-κB may have self-regulating activity [29-32]. Therefore, troxerutin modulates the miRNA-146a/NF-κB inflammatory signaling pathway to play as a regulatory negative feedback loop and reduce the inflammatory outcomes of diabetes in aortic tissue (Fig. 7). It seems that further studies with direct manipulation of miRNA-146a expression and its methylation are required to find an association between this miRNA and beneficial effects of troxerutin on endothelial cells and inflammatory responses as well as oxidative stress in diabetic animals. Since the oxidative stress and inflammatory reactions have crucial reciprocal effects to boost each other, it is better to investigate the anti-inflammatory and anti-oxidative effects of troxerutin in parallel in diabetic vessels and the cross-talk of these mechanisms through interaction with miRNA-146a.
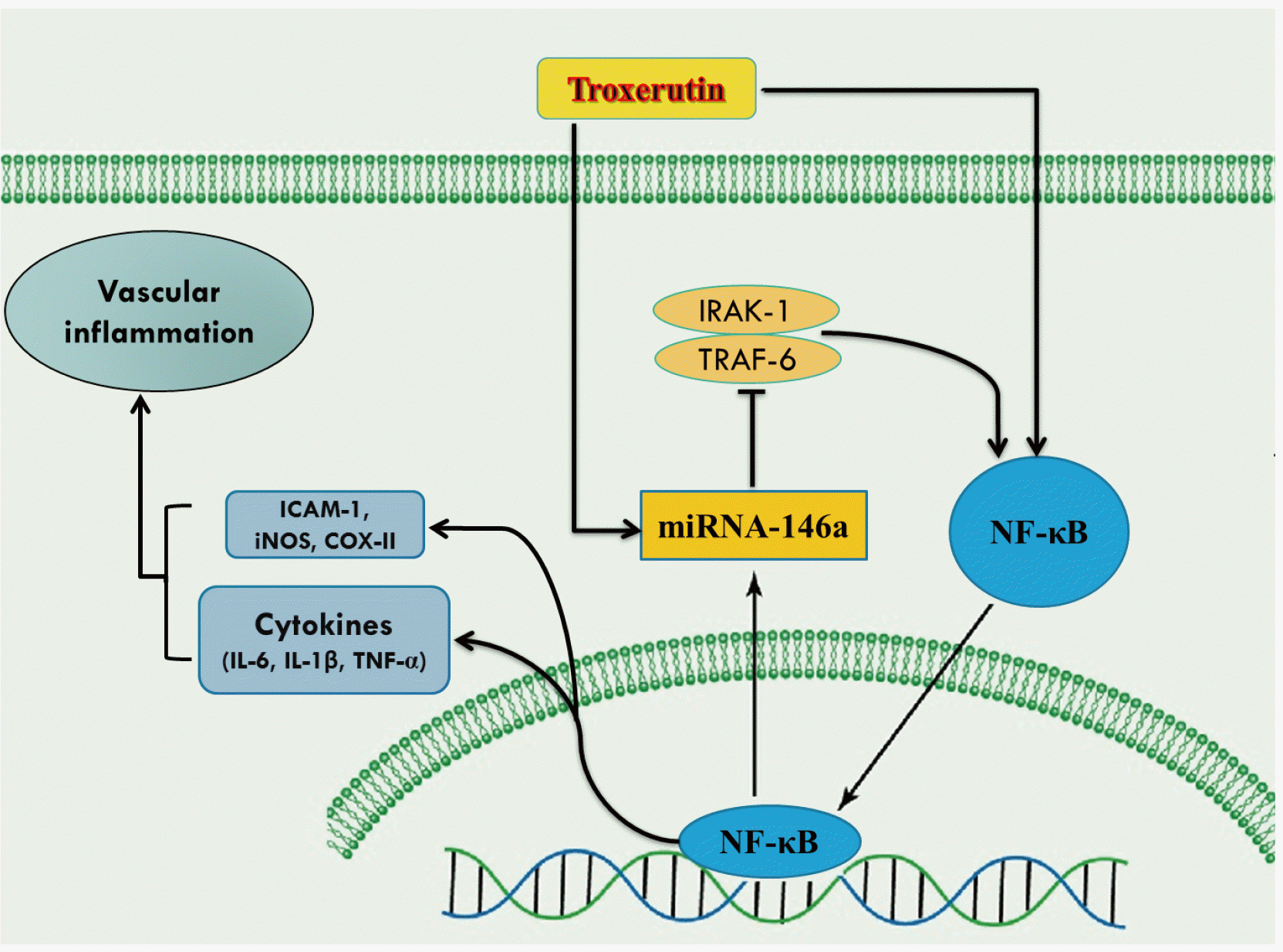
In conclusion, troxerutin therapy alleviated the diabetic-induced overproduction of inflammatory cytokines, as well as the endothelial adhesion molecule and inducible enzymes via regulating miRNAs-NF-κB signaling pathway in the aorta of STZ-induced diabetic rat (Fig. 7). The present results imply the protective effects of troxerutin on vascular complications of type-I diabetes.
Go to :
 XML Download
XML Download