

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Glioblastoma (GBM) is the most common and aggressive brain tumor and constitutes approximately 54% of all gliomas, frequently infiltrating into surrounding brain tissues owing to its high migration and invasion characteristics [1,2]. Despite advances in diagnostics and clinical therapeutic regimens, the median survival for GBM patients is approximately 15 months [3,4]. Therefore, there is a critical need to understand the molecular mechanisms of glioma cell migration and invasion to guide the development of new therapeutic strategies.
Substantial amounts of data have confirmed that the development of tumors is related to the abnormal localization and dysfunction of many cellular proteins [5,6]. XPO1/chromosomal region maintenance 1 (CRM1) plays a prominent role in the regulation of nuclear protein export [7], and overexpression of CRM1 has been reported in pancreatic, esophageal, osteosarcoma, ovarian, and gastric tumors, as well as in gliomas [8-13]. Thus, the nuclear exporter protein CRM1 may act as a novel potential therapeutic target for these deadly diseases. A large number of CRM1 inhibitors were discovered. Previous reports have shown that CRM1 specific inhibitors (such as Laptomycin B [LMB], KPT-185, -330 and so on) modify CRM1 at the reactive cysteine 528 site and abrogate the interaction between CRM1 and its cargo protein [14,15]. However, most of the currently available CRM1 inhibitors irreversibly bind to CRM1 and exhibit a relatively unfavorable drug tolerability profile [16]. Accordingly, a safer, more effective, and less toxic CRM1 inhibitor is urgently needed. Our previous study showed that the novel reversible CRM1 inhibitor S109 suppressed the proliferation of glioma cells both in vitro and in vivo [12]. However, the role of CRM1 in the migration and invasion of tumor cells remains unclear.
The signal transducer and activator of transcription 3 (STAT3) is a transcription factor that has been identified in various types of tumors, including breast, pancreatic, lung, prostatic, and brain cancers [17,18]. Activation of STAT3 is thought to play an important role in several features of malignant neoplasia including tumor cell survival, metastasis, and resistance to chemotherapy [19-22]. Evidence strongly suggests that STAT3 is an important oncogenic driver of GBM, and it has been shown that STAT3 inhibition in glioma can decrease the capacity for cell migration and invasion through reducing the activity and expression of MMP2 [23]. Moreover, siRNA-mediated silencing of STAT3 identifiably suppressed the chemo-responsiveness and migratory ability of glioma stem cells, and STAT3 plays an important role in maintaining self-renewal of glioma stem cells [24]. Given the apparent role of STAT3 in the genesis and progression of glioma, inactivation of the STAT3 signaling pathway may be an effective treatment strategy for these lethal diseases.
In this study, we investigated the effects of the CRM1 inhibitor S109 on migration and invasion of glioma cells. Results showed that S109 suppressed the invasion and migration of glioma cells partly due to the inactivation of the STAT3/MMP2 signaling pathway. Furthermore, our study provides insights into the applicability of using S109 as a potential targeted drug in gliomas.
METHODS
Cell culture and reagents
The human glioma cell line U251 was purchased from the Shanghai Cell Bank, Chinese Academy of Sciences. U87 cells, glioblastoma of unknown origin (catalog number: ATCC HTB-14), were derived from ATCC. These cells were cultured in DMEM supplemented with 10% FBS. These cell lines were grown in a humidified incubator containing 5% CO2 at 37°C. Primary antibodies against CRM1 (sc-74454) and actin (sc-58673) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The antibodies to STAT3 (9139s), p-STAT3 (9145s), and MMP2 (13132s) were purchased from Cell Signaling Technology (Beverly, MA, USA). The S109 compound was synthesized by the Suzhou Komanda Drug Development Company. S109 was dissolved in DMSO to create a 10 mM solution, which was diluted to different concentrations of medium before use.
Wound-healing assay
The migration behavior of glioma cells was evaluated using the wound-healing assay, according to our previous report [25,26]. U87 and U251 cells were seeded in 6-well plates and allowed to attach overnight. A rectangular lesion was created by using a plastic pipette tip, and cells were then incubated in serum-free media. The cells were incubated in serum-free media and treated with either 0.1% DMSO or S109. After incubation for 24 h or 48 h, cell migration into the wounded areas was observed and photographed using an inverted microscope. The experiments were independently performed three times.
Transwell invasion assay
Cell invasion assay was performed using a transwell system as described previously [27,28]. Culture inserts were coated with Matrigel and placed into the wells of 24-well culture plates. U87 and U251 cells were treated with either 0.1% DMSO or S109 in serum-free media and added to the top chamber. In the lower chamber, DMEM media containing 10% FBS was added. After 30 h of incubation, the noninvasive cells were removed from the upper chamber, the filters were fixed in 4% methanol for 20 min, and then stained with a 0.1% crystal violet solution for 30 min. The invading cells on the filter were counted from six randomly selected fields. The experiments were performed at least three times.
Western blotting
U87 and U251 cells were treated with variable concentrations of S109. The supernatants were collected by centrifugation at 13,000 g for 30 min and stored at –20°C. The total protein extracts from treated or untreated cells were used to western blot analysis within three days as described elsewhere [29-31]. The expression patterns of STAT3, p-STAT3, MMP2 were detected using specific antibodies, and β-actin were used as the loading control (all diluted to 1:1,000).
Gelatin zymography assay
The activity of MMP2 was assessed by gelatin zymography assay. The cells were seeded in 12-well culture plates and cultured for 24 h at 37°C. The cells were washed twice with PBS and incubated for an additional 24 h in serum-free medium supplemented with different concentrations of S109. Then, in the SDS loading buffer, media was harvested, centrifuged and resuspended without the use of β-mercaptoethanol. All samples were analyzed by 10% SDS-PAGE (containing 0.2% gelatin). Gels were washed in 2.5% Triton X-100 three times to remove SDS and then incubated overnight in reaction buffer. Subsequently, gels were stained with 0.25% Coomassie Brilliant blue R-250 and destained with 40% methanol and 10% acetic acid. The gelatinolytic activity of MMP2 in the gel was detected as clear white bands on a dark background.
Establishment of CRM1-wild-type (WT) and CRM1-C528S stable cell lines
Human CRM1 wild-type and C528S mutant genes were cloned into pWPXLD lentiviral expression vector containing a sequence coding for a flag tag. Then the constructed plasmids were sequenced. The viruses were produced in 293FT cells and used to infect U87 glioma cells according to our previous literature [12]. After 48 h infection, the cells were continuously cultured in medium containing 2.5 μg/ml puromycin. The surviving cells were cultured into cell lines stably expressing CRM1-WT or CRM1-C528S.
Statistical analysis
The statistical analyses were performed using the GraphPad Prism 5 software package (GraphPad Software, Inc., San Diego, CA, USA). Each experiment was repeated independently at least three times. Values are expressed as means ± standard error of the mean. The selected chart represents results from one of the repeated experiments. Comparisons of the mean values between the control and treated groups were performed using Students t-test. p-values < 0.05 were considered statistically significant.
RESULTS
Effects of S109 on the migration of glioma cells
To investigate the influence of S109 on the migration and invasion of glioma cells, wound-healing and transwell invasion assays were performed. Compared to control cells, the number of migratory U87 cells was decreased to 42.7%, 34.5%, and 18.8% at 24 h and 52.6%, 30.3%, and 15.3% at 48 h in response to treatment with 0.5 μM, 1 μM, and 2 μM S109, respectively (Fig. 1). Similar results were obtained in U251 cells. After 24 h incubation, the average number of migratory U251 cells was decreased to 61.9%, 46.9%, and 30.0%, and at 48 h these numbers were decreased to 61.5%, 49.7%, and 32.7% following treatment with 0.5 μM, 1 μM, and 2 μM S109, respectively. To exclude the possibility that the concentrations of S109 used in the migration assay had no obvious cytotoxic effect on U87 and U251 cells, we used the CCK8 assay to assess the effect of S109 (0–2 μM) on cell viability after 24 h or 48 h. We found that the lower doses of S109 (0–2 μM) had no obvious effect on cell viability after 24 h and 48 h (Supplementary Fig. 1, Table 1). Our results demonstrated that S109 significantly inhibited the migration of glioma cells.
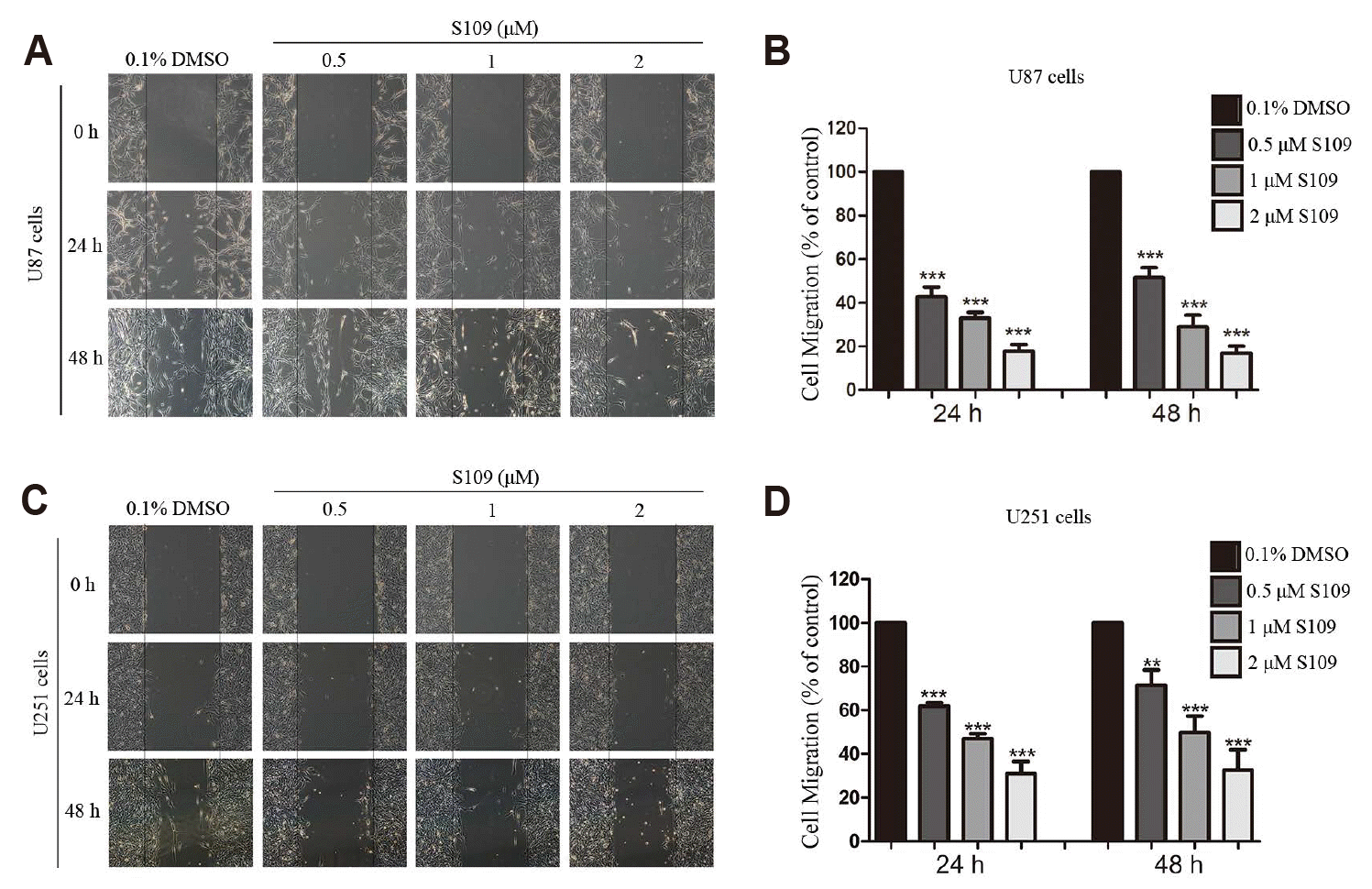
Fig. 1
S109 suppressed the migration of U87 and U251 cells.
(A, C) U87 and U251 cells were treated with vehicle or various concentrations of S109 for 24 h and 48 h. Wound-healing assay was then performed to evaluate cell migratory capacity. (B, D) Quantitative analysis of migratory cell numbers. The number of migratory cells was counted and then normalized to that of the control group. The data are presented as the means ± standard error of the mean from three independent experiments, **p < 0.01, ***p < 0.001.
![]()
Effects of S109 on the invasion of glioma cells
Next, we further investigated the effects of S109 on human glioma cell invasion via transwell invasion assay. The results showed that S109 induced a dose-dependent reduction in invasive cell numbers with increasing concentrations of S109. Compared with the control group, the mean invasive U87 cell number was reduced to 60.2%, 49.3%, and 13% at 0.25 μM, 0.5 μM, and 1 μM S109 treatment, respectively. Similarly, the exposure of U251 cells to 0.25 μM, 0.5 μM, and 1 μM S109 reduced invasion rate to 64.8%, 49.3%, and 16.0%, respectively (Fig. 2). Taken together, these data imply that S109 effectively inhibited the migration and invasion of glioma cells.
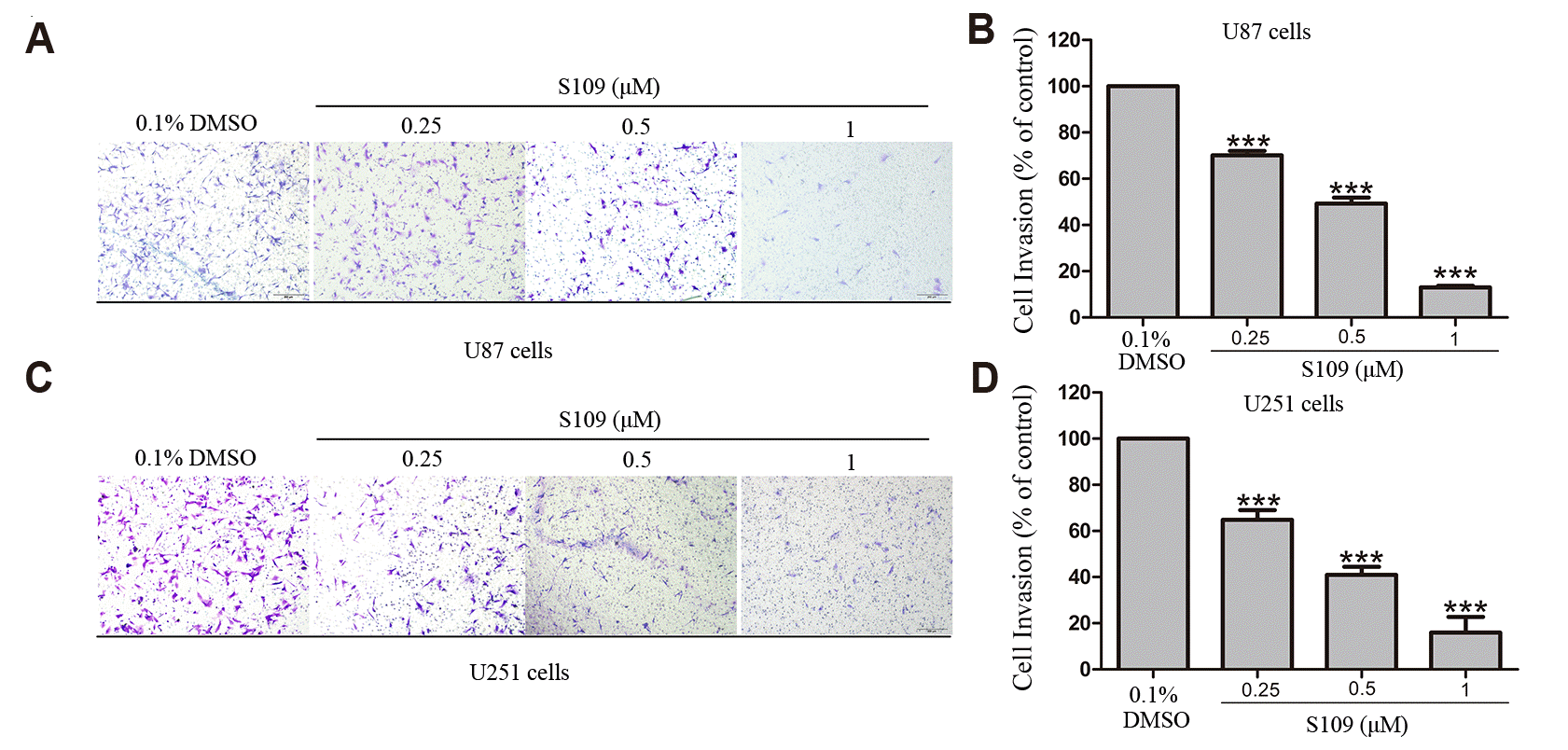
Fig. 2
S109 treatment decreased the invasive ability of U87 and U251 cells.
(A, C) U87 and U251 cells were treated with vehicle or S109 (0.25 μM, 0.5 μM, and 1.0 μM) for 30 h, and then cell invasion was assessed by transwell invasion assay. The invasive cells were stained with a 0.1% crystal violet solution for 30 min. (B, D) Quantitation of numbers of cells that invaded through the filter. The number of invading cells was counted and normalized to the control group. The data are expressed as the means ± standard error of the mean from three independent experiments, ***p < 0.001.
![]()
S109 inhibits the expression and activity of MMP2 by blocking STAT3 signaling
MMP2 and MMP9 are vital enzymes involved in the degradation of the extracellular matrix (ECM) and the components of the basement membrane [32]. Given that the activity of MMP2 is considered to be a vital factor in tumor invasion, we first evaluated the effects of S109 on the activity and expression of MMP2 in U87 and U251 cells via gelatin zymography assay and Western blot analysis, respectively. As illustrated in Fig. 3A and B, S109 reduced the activity of MMP2 in U87 and U251 cells in a dose-dependent manner. Next, we further evaluated the expression of MMP2 after treatment with S109. The data indicated that treatment with increasing concentrations of S109 induced a significant decrease in the expression level of MMP2 in U87 and U251 cells (Fig. 3C–F).
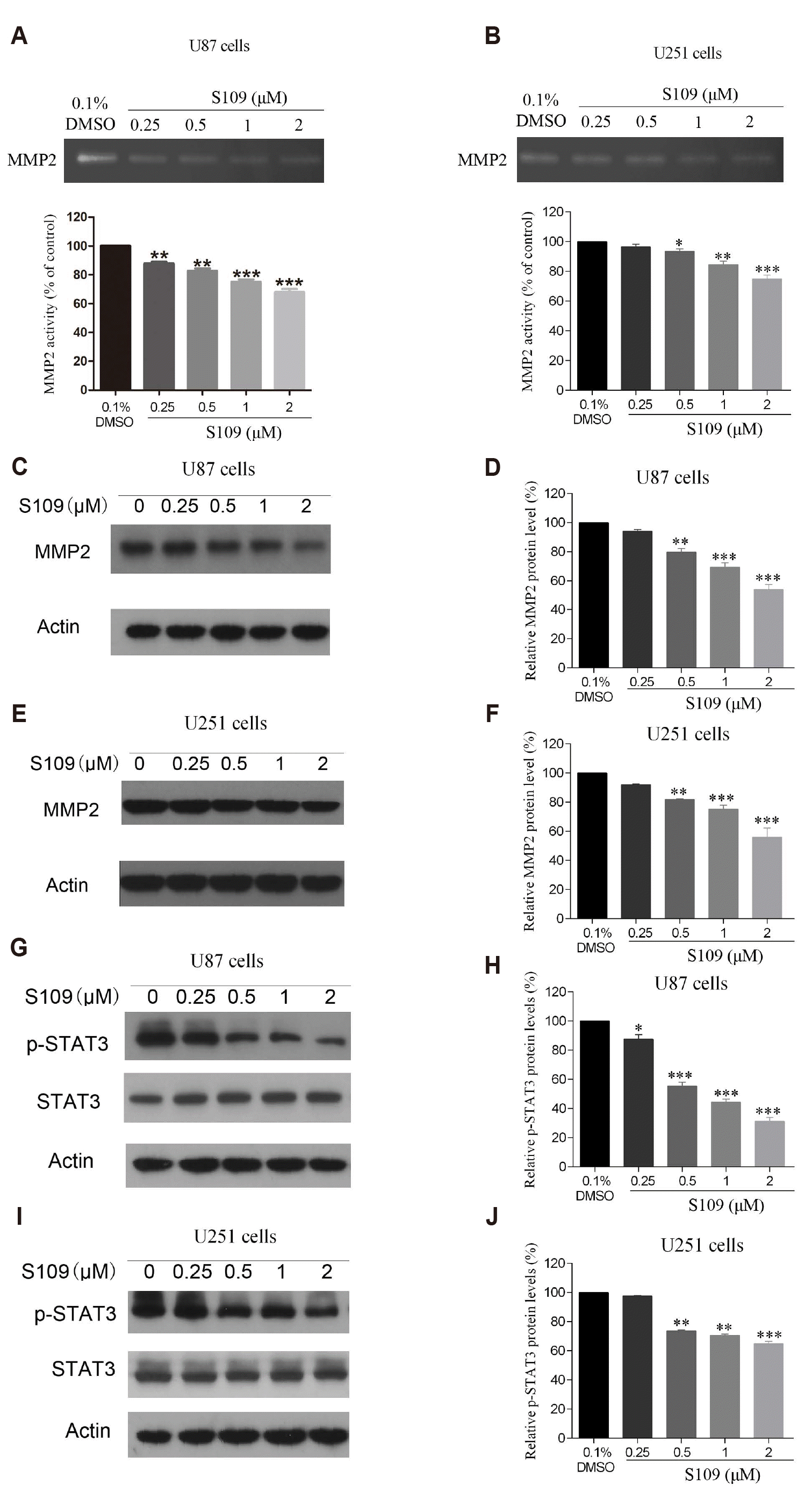
Fig. 3
S109 inhibited the activity and expression of MMP2 via inactivation of STAT3 signaling.
(A, B) Representative picture and quantitative MMP2 activity of gelatin zymography assay. Cells were treated with S109 at the indicated concentrations, and then the gelatin zymography assay was performed. (C–F) The effects of S109 on MMP2 expression by Western blot analysis. (G–J) Phosphorylated STAT3 and total STAT3 expression were examined in U87 and U251 cells treated with S109. All data presented here are the means ± standard error of the mean from three independent experiments, *p < 0.05, **p < 0.01, ***p < 0.001.
![]()
Previous studies have indicated that MMP2 gene expression is transcriptionally regulated by various transcription factors, including STAT3 [33]. The activity of STAT3 is necessary for invasion in gliomas. Down-regulation of STAT3 reduces invasive activity via suppressing the expression of MMP2 in gliomas [34,35]. Hence, we further investigated the effects of S109 on the activity and expression of STAT3 in U87 and U251 cells, and the results are shown in Fig. 3G and I. The level of phosphorylated STAT3 but not the total STAT3 was markedly decreased in a dose-dependent manner after treatment with S109. The above data suggested that S109 might inhibit the expression and activity of MMP2 by inactivating STAT3 signaling.
The CRM1 Cys528 mutation abrogates the inhibition of glioma cell migration and invasion by S109
The Cys528 residue in the cargo-binding pocket of CRM1 is essential for the inhibitory effect of the CRM1 inhibitor [36]. Our previous study demonstrated that S109 possesses potent anti-proliferative activity through selectively binding to the Cys528 residue of CRM1 in gliomas [12]. In the present study, previously constructed U87 cell lines stably expressing CRM1-WT and CRM1-C528S were used to further investigate whether S109 inhibits the invasion and migration of glioma cells by specifically inhibiting CRM1 [12]. The identification of CRM1-WT and CRM1-C528S stable cell lines is shown in Supplementary Fig. 2. The results of wound-healing and transwell invasion assays suggested that S109 treatment markedly attenuated the migratory and invasive ability of glioma cells that express the wild-type CRM1. However, the inhibitory effect of S109 on the migration and invasion of glioma cells was nearly abolished in the cells expressing the Cys528 mutant CRM1 (Fig. 4A–D). Subsequently, we further examined the effect of S109 on p-STAT3 expression in CRM1-WT and CRM1-C528S expressing cells. We found that S109 inhibited the phosphorylation of STAT3. In contrast, the CRM1 Cys528 mutation abrogated the inhibition of p-STAT3 expression induced by S109 treatment. The results are shown in Fig. 4E. These findings further provide evidence suggesting that S109 can inhibit glioma cell migration and invasion through selectively binding the Cys528 residue of CRM1.
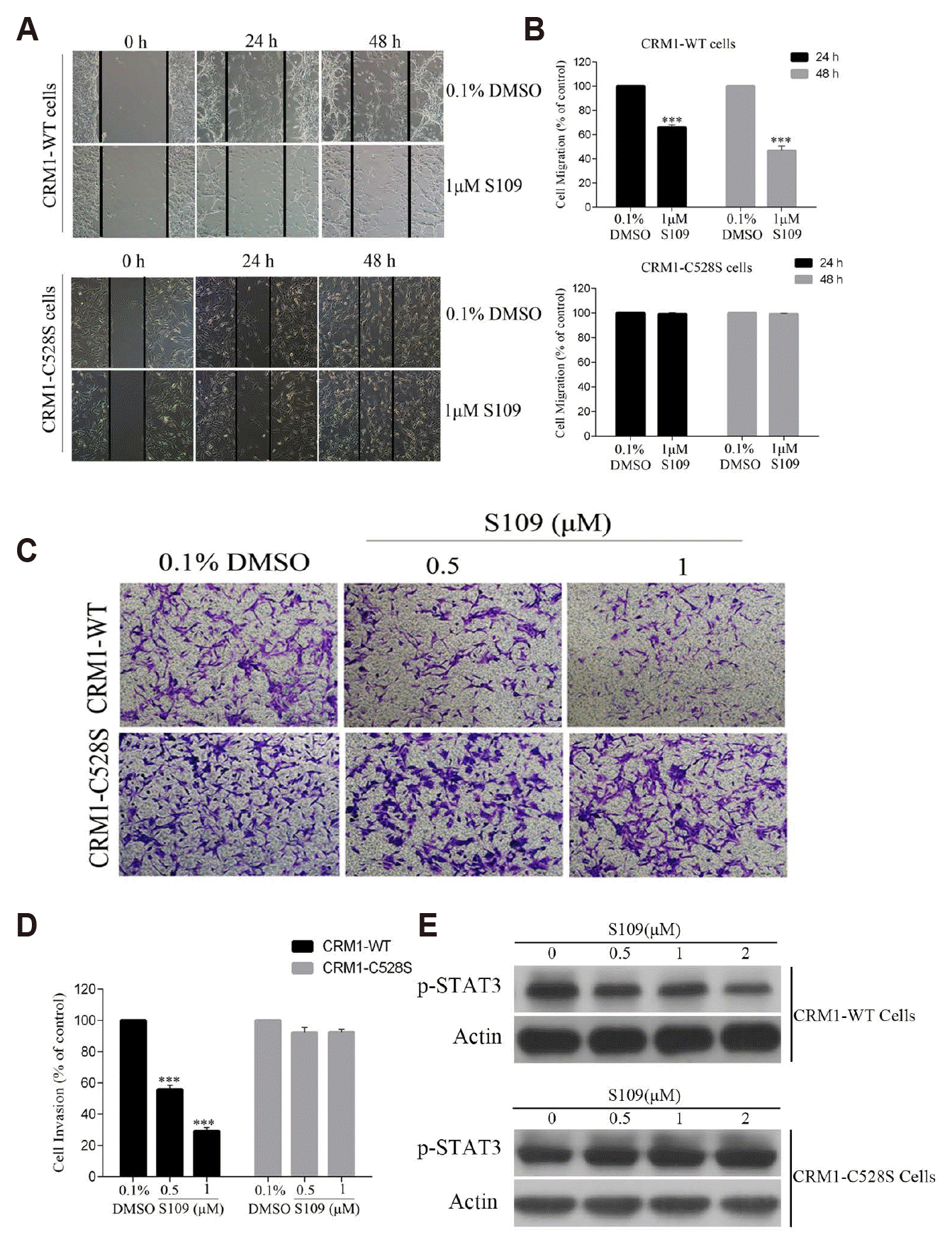
Fig. 4
The mutation of chromosomal region maintenance 1 (CRM1) C528 abolished the anti-migration and anti-invasion activity of S109 in glioma cells.
U87 cells expressing wild-type (WT) and C528S-mutant CRM1 were treated with S109, and then migration and invasion were assessed by wound-healing cell migration assay (A, B) and transwell invasion assay (C, D). Results are expressed as means ± standard error of the mean of three independent experiments. ***p < 0.001 compared with the control group. The invasive cells were stained with a 0.1% crystal violet solution for 30 min. (E) The effect of S109 treatment on p-STAT3 expression level in CRM1-WT and CRM1-C528S cells. U87, U87-CRM1-WT and U87-CRM1-C528S cells were treated with S109 (0–2 μM) for 24 h respectively, and then the total protein was exacted. The expression level of p-STAT3 were examined by Western blot assays.
![]()
DISCUSSION
GBM is the most common and the most devastating primary brain tumor [1], and the high recurrence rate and mortality are attributed to the highly invasive nature of these tumor cells [37]. Therefore, pharmacological suppression of glioma cell migration and invasion may be deemed a promising strategy for glioma treatment. Here, we found that the CRM1 inhibitor S109 effectively inhibited the migration and invasion of human glioma cells in vitro. Furthermore, our data suggested that S109 exerts its anti-migrative and anti-invasive activities through inactivation of the STAT3/MMP2 pathway. Overexpression of CRM1 has been identified in several types of malignant tumors and is associated with aggressive behavior and poor survival [38]. Therefore, CRM1 is considered a promising therapeutic target for gliomas. Our previous study demonstrated that the CRM1 inhibitor S109 significantly suppressed the proliferation of glioma cells both in vitro and in vivo and prolonged the survival of nude mice [12]. In the present study, we found that S109 treatment significantly decreased the migrative and invasive ability of glioma cells. Previous reports have shown that CRM1 can bind to a hydrophobic nuclear export signal (NES) peptide of cargo substrate. The hydrophobic NES of the cargo protein binds to a hydrophobic groove of CRM1 containing an active site Cys528. CRM1 inhibitors bind to the active site Cys528 in CRM1 and site-directed mutagenesis of Cys528 to Ser528 will prevent binding of CRM1 by LMB and other CRM1 inhibitors [15,39]. In our study, we also found that mutation of CRM1 abolished the anti-invasive effect of S109. These data imply that CRM1 also plays an important role in the migration and invasion of glioma cells.
The migration of tumor cells across tissue barriers demands the degradation of specific components of the extracellular matrix [40]. ECM components confer cell migrative and invasive properties, prevent apoptosis and senescence, and contribute to immunosuppression [41]. MMP2 is a member of the extracellular matrix-degrading enzyme family, which is closely related to the invasion and migration of tumors, and plays an important role in matrix remodeling, angiogenesis, invasion, and tumor metastasis [42]. MMP2 is considered a diagnostic and prognostic biomarker and is also a potential therapeutic target [43]. CRM1-selective inhibitors can reduce migratory and invasive properties via suppressing the secretion and expression of MMP2 in prostate cancer cells [44]. Consistently, we found that S109 treatment resulted in a significant reduction in the levels of MMP2 protein. Further investigation demonstrated that S109 inhibited the activity of MMP2 in the U87 and U251 cells. These results suggest that S109 induced the inhibition of glioma cell migration and invasion by decreasing the activity and expression of MMP2.
STAT3 is constitutively activated in various cancers and is phosphorylated in the cytoplasm by growth factor receptors, Janus kinase, and Src kinase families [45,46]. The phosphorylated STAT3 forms homodimers and transfers into the nucleus, thereby regulating the transcription of target genes such as MMP2 [47]. Previous reports have suggested that STAT3 is persistently activated through constant nucleocytoplasmic shuttling, which is regulated by CRM1 [48,49]. STAT3 contains several NES elements through which it binds to CRM1 for nuclear export [49,50]. Inhibition of CRM1-mediated nuclear export can slow down the nucleocytoplasmic shuttling of activated STAT3 resulting in reduced tyrosine phosphorylation, decreased induction of STAT3 target genes, and increased apoptosis [51]. Previous studies have indicated that the STAT3/MMP2-involved pathway is significant to cancer invasion and metastasis [52]. In addition, STAT3 can promote the transcription of the target gene MMP2 via direct interaction with a consensus site in the MMP2 promoter [47]. Down-regulation of STAT3 phosphorylation leads to decreased MMP2 activity, migration, and invasion in human brain cancer cells [35]. Consistent with these studies, our results showed that S109 treatment decreased the level of STAT3 phosphorylation. Taken together, our data support that CRM1 inhibition by S109 decreases phosphorylation of STAT3, which in turn inhibits the expression and activity of MMP2. Ultimately, these processes lead to suppression of migration and invasion in glioma cells.
In summary, we elucidated that a novel CRM1 selective reversible inhibitor, S109, inhibited glioma cell migration and invasion by inactivation of the STAT3/MMP2 signaling pathway. However, further in-depth studies are needed to confirm how S109 regulates STAT3 activity. Based on the evidence provided here, our data suggest that S109 may act as an effective chemotherapeutic drug for the treatment of gliomas.
 XML Download
XML Download