

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Signal transducers and activators of transcription (STATs) are transcription factors that locate in the cytoplasm, where they are phosphorylated by membrane-bound receptors and then translocate to the nucleus [1]. STATs control the entire hematopoietic process by regulating cytokine binding to specific cell-surface receptors [1]. STAT4, in particular, is a member of the STAT family, which is activated by interleukin-12 (IL-12) through its receptor and plays an essential downstream role in cell differentiation and proliferation. The system of type I interferon (IFN) plays an important role in the initiation and development of systemic lupus erythematosus (SLE) [2], and STAT4 induces the output of IFN-ÿ. STAT4 is also involved in the development of Th17 cells which have a dominant role in autoimmune-associated inflammation [3]. The polymorphism of STAT4 rs7574865 is located within the third intron of the gene. Notably, there is no evidence to suggest that this intronic polymorphism by itself is directly related to changes in protein function. Nevertheless, the STAT4 rs7574865 polymorphism shows the strongest association with autoimmunity, and several SNPs are strongly associated with this intronic SNP [4]. The STAT4 rs7574865 polymorphism has been reported to be associated with SLE in multiple ethnic groups [5-28]. Allelic frequencies of genes are usually significantly different among populations; therefore, ethnicity-specific association studies are needed to confirm genetic associations in various ethnic populations. In this research, we used a meta-analysis method to examine the relationship between the STAT4 rs7574865 polymorphism and SLE susceptibility in various populations.
Go to : 
MATERIALS AND METHODS
Identification of eligible studies and data extraction
We searched for reports investigating the relationships between SLE and STAT4 polymorphisms. The MEDLINE and EMBASE databases were searched for available studies which analyzed STAT4 polymorphisms in SLE patients (up to January 2020). As Medical Subject Headings (MeSH) or text words, keyword combinations such as “interferon regulatory factor,” “STAT4,” “polymorphism,” “rs7574865,” “systemic lupus erythematosus,” and “SLE” were entered. References were also investigated in the selected studies to identify additional research not indexed by the electronic databases. No limitations were imposed on language or region. Studies were included if: (1) case-control studies, (2) data were original (independent of studies), and (3) adequate data were given to measure an odds ratio (OR). We excluded studies according to the following criteria: (1) studies containing duplicate data, (2) studies in which they were not possible to determine the number of null and wild genotypes or alleles, and (3) studies in which family members were studied, because these analyzes are based on considerations of linkage. From each research, the following information was extracted: author, year of publication, ethnicity of the study population, demographics, number of cases and controls, and the allele frequency of the STAT4 rs7574865 poly-morphism. Any differences among reviewers were resolved by consensus, and the meta-analysis was carried out in accordance with the guidelines of PRISMA [29].
Evaluation of statistical associations
In each study the frequency of the STAT4 rs7574865 T allele was determined using the method of allele counting. Meta-analyzes were then carried out using the contrast model for the alleles. For each report, point estimates of risk, ORs, and confidence intervals (CI) of 95% were determined. Moreover, heterogeneities within and between the studies were evaluated using Q-statistics from Cochran. This heterogeneity test evaluated the null hypothesis all studies were evaluating the same effect. The effect of heterogeneity was measured using I2, which ranged from 0%∼100% and reflected the proportion of inter-study variation that could be due to heterogeneity rather than chance [30]. Nominally allocated as low moderate, and high values were I2 values of 25%, 50%, and 75%. The model of fixed effects presumed that genetic factors had similar effects on SLE susceptibility throughout all studies, and that the observed differences between studies were caused by chance alone [31]. The model of random effects assumed that different studies displayed significant variability and measured both in-study sampling error and inter-study variance [32]. In case of significant inter-study heterogeneity, the random effects model was used [33]. Otherwise, fixed effect model was chosen. Statistical manipulations were carried out using the Comprehensive Meta-Analysis software (Biostat, Englewood, NJ, USA).
Evaluation of heterogeneity, sensitivity, and publication bias
Subgroup analyzes were conducted based on ethnicity in order to explore the potential sources of heterogeneity found in this meta-analysis. In addition, sensitivity analysis was carried out not only to determine the effect of each individual study on pooled ORs by removing each individual study, but also to examine the statistically robust effects of this meta-analysis. While funnel plots have been used to detect publishing bias, they demand several studies of different sample sizes involving subjective judgements. Therefore, we evaluated publishing bias using Egger's linear regression test, which measures funnel plot asymmetry using a natural OR logarithm scale [34].
Go to :
RESULTS
Studies included in the meta-analysis
A total of 259 studies were identified using electronic and manual searches, of which 36 were chosen for a full-text review based on the contents of the title and abstract. Twelve of these 36 studies have been omitted, either because they did not contain data on rs7574865 or overlapping data. Of the 24 studies that met the inclusion criteria [6-29], 3 studies contained data from 3 different groups [10,24,28], 1 studies contained data from 5 different groups [22] and 1 studies contained data from 2 different groups [20]. In this meta-analysis, which contained 22,898 SLE patients and 24,838 controls from 11 European, 17 Asian, 4 Latin American, 21 Arab, and 2 African American populations, a total of 36 different comparisons were considered (Table 1). Meta-analyzes specific to ethnicity were carried out on each ethnic population.
Table 1
Characteristics of individual studies included in meta-analysis
| Author | Country | Ethnicity | Number | T allele (%) | Association | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|||||||||
| SLE | Control | SLE | Control | OR | 95% CI | p-value | |||||
| Gupta et al., 2018 [5] | India | Asian | 394 | 583 | 32.9 | 26.2 | 1.382 | 1.134∼1.685 | 0.001 | ||
| Salmaninejad et al., 2017 [6] | Iran | Arab | 50 | 281 | 39.0 | 31.9 | 1.368 | 0.882∼2.123 | 0.162 | ||
| Beltrán Ramírez et al., 2016 [7] | Mexico | LA | 128 | 326 | 53.1 | 39.4 | 1.742 | 1.302∼2.331 | <0.001 | ||
| Ciccacci et al., 2014 [8] | Italy | European | 239 | 278 | 35.0 | 20.0 | 2.153 | 1.626∼2.849 | <0.001 | ||
| Dang et al., 2014 [9] | China | Asian | 370 | 576 | 40.1 | 30.9 | 1.499 | 1.236∼1.818 | <0.001 | ||
| Chai et al.-1, 2014 [10] | Malaysia | Asian | 93 | 110 | 44.1 | 32.7 | 1.621 | 1.082∼2.427 | 0.019 | ||
| Chai et al.-2, 2014 [10] | China | Asian | 245 | 294 | 46.3 | 37.8 | 1.423 | 1.115∼1.815 | 0.005 | ||
| Chai et al.-3, 2014 [10] | India | Asian | 22 | 26 | 31.8 | 42.3 | 0.636 | 0.275∼1.474 | 0.291 | ||
| Kadota et al., 2013 [11] | Japan | Asian | 75 | 190 | 47.3 | 33.4 | 1.790 | 1.218∼2.631 | 0.003 | ||
| Mirkazemi et al., 2013 [12] | Iran | Arab | 280 | 281 | 41.3 | 31.9 | 1.502 | 1.176∼1.918 | 0.001 | ||
| Sanchez et al.-1, 2011 [13] | USA | AF | 1,569 | 1,893 | 16.8 | 14.5 | 1.190 | 1.045∼1.355 | 0.009 | ||
| Sanchez et al.-2, 2011 [13] | USA | AF | 155 | 131 | 15.0 | 10.7 | 1.493 | 0.906∼2.462 | 0.116 | ||
| Li et al., 2011 [14] | China | Asian | 748 | 554 | 41.0 | 43.2 | 0.914 | 0.781∼1.070 | 0.263 | ||
| Koga et al., 2011 [15] | Japan | Asian | 282 | 222 | 46.3 | 34.7 | 1.622 | 1.256∼2.096 | <0.001 | ||
| Sanchez et al., 2010 [16] | USA | LA | 727 | 595 | 39.1 | 33.9 | 1.247 | 1.063∼1.463 | 0.007 | ||
| Kiyohara et al., 2009 [17] | Japan | Asian | 152 | 427 | 37.5 | 29.0 | 1.466 | 1.113∼1.931 | 0.006 | ||
| Suarez-Gestal et al., 2009 [18] | Spain | European | 960 | 661 | 33.1 | 24.0 | 1.570 | 1.341∼1.839 | <0.001 | ||
| Namjou et al., 2009 [19] | Korea | Asian | 661 | 781 | 33.1 | 24.0 | 1.570 | 1.341∼1.839 | <0.001 | ||
| Yang et al.-1, 2009 [20] | China | Asian | 910 | 1440 | 46.1 | 33.4 | 1.705 | 1.512∼1.923 | <0.001 | ||
| Yang et al.-2, 2009 [20] | Thai | Asian | 278 | 383 | 48.7 | 35.2 | 1.747 | 1.398∼2.183 | <0.001 | ||
| Han et al., 2009 [21] | China | Asian | 8,255 | 4,199 | 42.3 | 32.7 | 1.505 | 1.426∼1.589 | <0.001 | ||
| Abelson et al.-1, 2009 [22] | Germany | European | 247 | 220 | 26.5 | 21.0 | 1.365 | 1.007∼1.851 | 0.045 | ||
| Abelson et al.-2, 2009 [22] | Italy | European | 221 | 207 | 39.4 | 19.3 | 2.711 | 1.989∼3.695 | <0.001 | ||
| Abelson et al.-3, 2009 [22] | Spain | European | 390 | 620 | 32.7 | 22.1 | 1.712 | 1.401∼2.094 | <0.001 | ||
| Abelson et al.-4, 2009 [22] | Argentina | LA | 171 | 171 | 41.5 | 31.9 | 1.518 | 1.110∼2.076 | 0.009 | ||
| Abelson et al.-5, 2009 [22] | Mexico | LA | 552 | 633 | 54.7 | 36.4 | 2.109 | 1.789∼2.487 | <0.001 | ||
| Kawasaki et al., 2008 [23] | Japan | Asian | 308 | 306 | 46.3 | 33.5 | 1.709 | 1.357∼2.153 | <0.001 | ||
| Kobayashi et al.-1, 2008 [24] | Japan | Asian | 227 | 745 | 43.8 | 31.1 | 1.726 | 1.391∼2.140 | <0.001 | ||
| Kobayashi et al.-2, 2008 [24] | Japan | Asian | 183 | 938 | 38.5 | 31.0 | 1.397 | 1.107∼1.762 | 0.005 | ||
| Kobayashi et al.-3, 2008 [24] | Japan | Asian | 165 | 212 | 43.9 | 30.4 | 1.792 | 1.328∼2.420 | <0.001 | ||
| Taylor et al., 2008 [25] | USA | European | 1,938 | 2,560 | 31.1 | 22.1 | 1.589 | 1.446∼1.747 | <0.001 | ||
| Palomino-Morales et al., 2008 [26] | Spain | European | 144 | 410 | 42.7 | 31.5 | 1.624 | 1.232∼2.140 | 0.001 | ||
| Harley et al., 2008 [27] | USA | European | 720 | 2337 | 32.0 | 23.0 | 1.576 | 1.384∼1.795 | <0.001 | ||
| Remmers et al.-1, 2007 [28] | USA | European | 575 | 416 | 31.0 | 22.0 | 1.597 | 1.299∼1.962 | <0.001 | ||
| Remmers et al.-2, 2007 [28] | USA | European | 349 | 416 | 31.0 | 23.0 | 1.504 | 1.198∼1.889 | <0.001 | ||
| Remmers et al.-3, 2007 [28] | USA | European | 115 | 416 | 29.0 | 22.0 | 1.453 | 1.046∼2.018 | 0.026 | ||
| Total | 24,838 | 22,898 | 36.8 | 28.5 | 1.549 | 1.459∼1.644 | <0.001 | ||||
![]()
Frequency of the STAT4 rs7574865 T allele in different ethnic groups
Among the controls, the mean frequency of the STAT4 rs7574865 T allele was 28.5%, varying from 14.3% to 35.7%. While the African American population showed the lowest prevalence of T alleles compared to other ethnic groups, the highest frequency was demonstrated by the Latin American population (Table 2). The T allele frequencies in the African American, European, Arab, Asian, and Latin American populations were 14.3%, 22.8%, 31.9%, 32.4%, and 35.7%, respectively, with respect to normal controls.
Table 2
Prevalence of the STAT4 rs7574865 polymorphism T allele
![]()
Association between the STAT4 rs7574865 polymorphism and SLE
Meta-analysis showed an association in all subjects between SLE and STAT4 rs7574865 polymorphism (OR= 1.549, 95% CI=1.459∼1.644, p<0.001) (Table 3, Figure 1). Analysis after ethnicity stratification revealed that in Europeans, Asians, Latin Americans, African Americans and Arabs, the STAT4 rs7574865 T allele was significantly associated with SLE (Table 3, Figure 2).
 | Figure 1Odd ratio and 95% confidence interval (CI) of individual studies and pooled data for the association between the STAT4 rs7574865 polymorphism T allele and systemic lupus erythematosus (SLE) in all subjects.
|
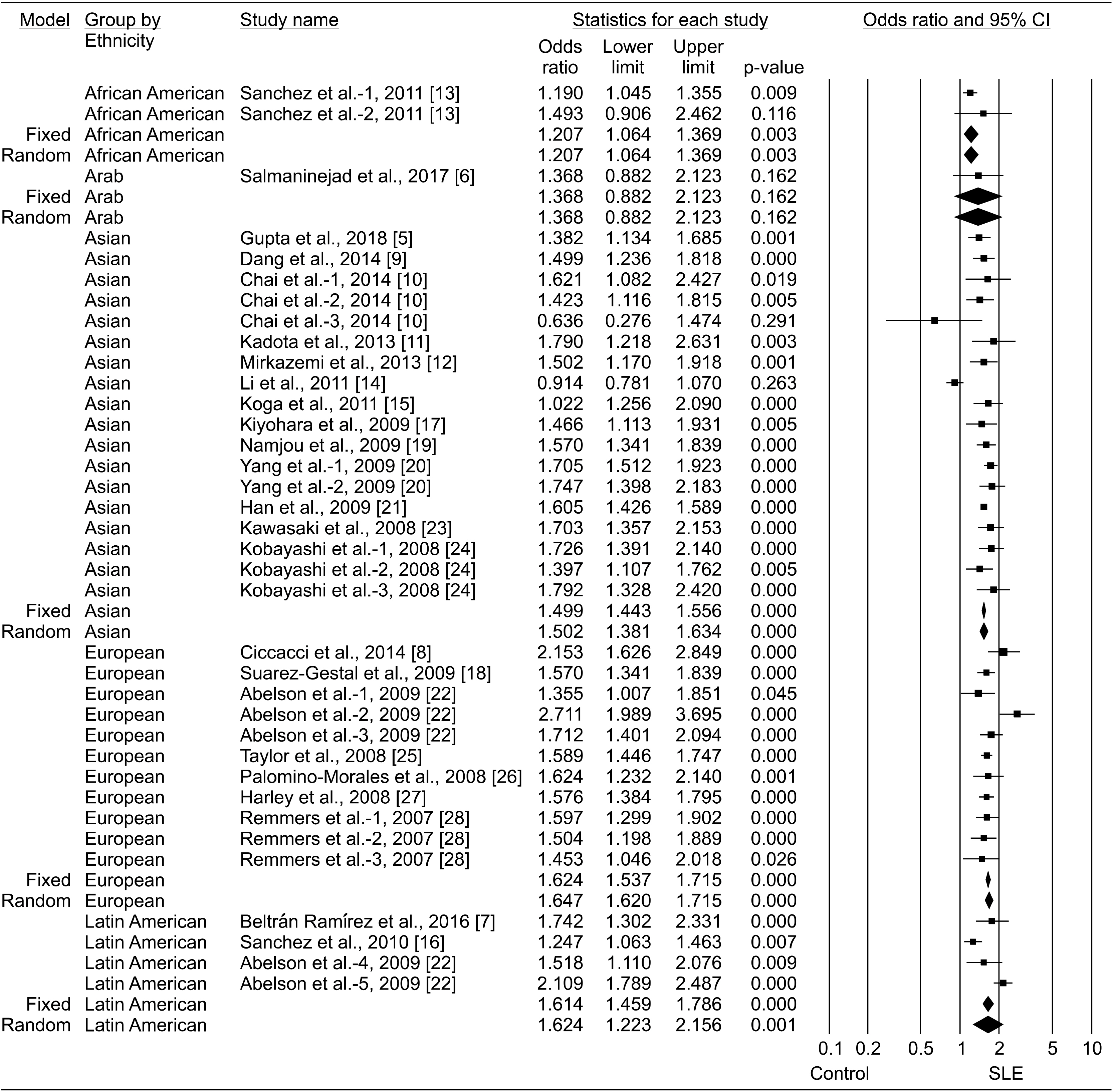 | Figure 2Odd ratios and 95% confidence intervals (CIs) of individual studies and of pooled data for the association between the T allele and systemic lupus erythematosus (SLE) in each ethnic group.
|
Table 3
Meta-analysis of the STAT4 rs7574865 polymorphism and SLE association
![]()
Association between the STAT4 rs7574865 polymorphism and clinical features of SLE
STAT4 rs7574865 was more strongly associated with SLE characterized by double-stranded (ds) DNA antibodies, nephritis, and/or age at diagnosis, 30 years [23,25]. Each of 3 studies indicated a strong association between the rs7574865 polymorphism and the production of anti-Sm antibodies, pericarditis, hematologic disorders and photosensitivity [8,14,20]. However, one study did not detect any correlation between the polymorphism and the specific clinical manifestations of SLE, such as photosensitivity, malar rash, discoid rash, oral ulcer, arthritis, pleuritis, pericarditis, proteinuria, seizures, leucopenia, anti-ds DNA, and antinuclear antibody, although sample size could be the reason for the insignificance in the comparisons [12].
Heterogeneity, sensitivity, and publication bias
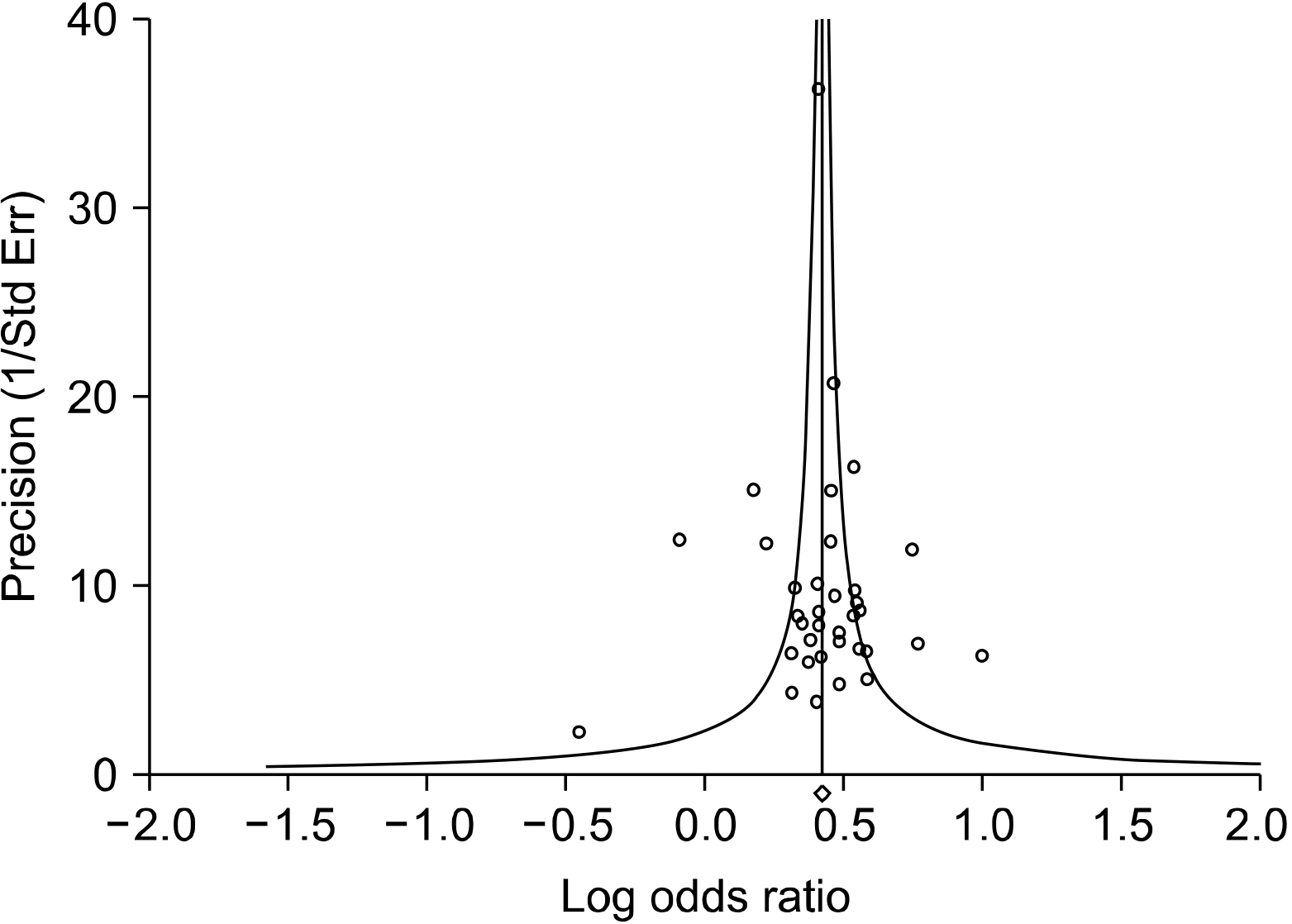
Meta-analysis showed between-study heterogeneity between STAT4 rs7574865 polymorphism and SLE in all study subjects and in the populations of Europe, Asia and Latin America; however, no heterogeneity was observed in the population of African America. Sensitivity analysis also showed that no single study significantly affected the pooled OR, and excluding two studies with potential overlap data [27,28] did not significantly affected the pooled OR (OR=1.551, 95% CI=1.450∼1.658, p< 0.001), thereby suggesting that the findings of this meta-analysis are robust. There was no proof of publication bias (Egger's regression test, p>0.1) (Table 3, Figure 3) in the funnel plot and Egger's regression test.
Go to :
DISCUSSION
In this study, through a comprehensive meta-analysis, we updated information regarding the association between the STAT4 rs7574865 polymorphism and SLE. The T allele prevalence was determined in various populations, and ethnic-specific meta-analyses were performed. Notably, rs7574865 T allele prevalence was found to differ among ethnic populations (14.3%∼35.7%). The mean T allele frequency in the controls was 28.5%, which was the lowest among African Americans and the highest among Latin Americans. In addition, ethnicity-specific meta-analysis revealed an association among European, Asian, Latin American, African-American Americans, and Arabs.
The result of this meta-analysis suggests that STAT4 could be a genetic risk factor for SLE, showing similar potential across multiple racial groups. Autoimmune diseases are triggered through the breakdown of self-tolerance; therefore, some common pathways are shared. STAT4 is involved in the signaling pathway of several essential cytokines, including IL-12, IFNs, and IL-23 [33]. Previously, STAT4-deficient mice had been shown to be resistant to animal models of diabetes, experimental autoimmune encephalomyelitis, and arthritis [35]. These results suggest that STAT4 could be a common genetic risk factor for autoimmune diseases.
Interestingly, increased expression of type I IFN-regulated genes has previously been documented in the peripheral blood leukocytes of SLE patients, suggesting the role of type I IFN in the pathogenesis of the disease [2]. Notably, rs7574836 polymorphism is located in the third intron of the STAT4 gene, and its functional significance has not yet been established. However, since the haplotype susceptibility is located within STAT4 intron 3, it may be responsible for splice variations or changes in STAT4's regulatory effects. In addition, it has also been shown that the STAT4 risk-haplotype increases the sensitivity of IFN-α signaling [36]. IFN-matured dendritic cells also present an increased risk of autoimmunity by supporting autoimmune T-and B-cells and by generating increased levels of anti-dsDNA antibody, both of which can contribute to the development of SLE [37].
This meta-analysis differs from an earlier meta-analysis of the relationship between STAT4 rs7574865 polymorphism and SLE risk conducted by Yuan et al. [38], because this study included 16 more studies, 14,445 more SLE patients, and 11,747 more controls in addition to meta-analysis conducted on Latin American and African American populations. The results of this meta-analysis showing the STAT4 rs7574865 polymorphism as a susceptibility factor for SLE in the European and Asian populations are in agreement with the results of the previous study. This meta-analysis also confirmed that the Latin Americans and African Americans associate the STAT4 rs7601754 polymorphism with SLE risk.
There are some limitations to this study which should be considered. First, the analysis may have been affected by heterogeneity and confounding factors, as is commonly found in any other meta-analysis. Second, the STAT4 polymorphism may be associated with clinical manifestations in addition to the susceptibility to disease. However, a thorough meta-analysis was not performed by the limited amount of data available on the association between the STAT4 rs7574865 polymorphism and severity of the disease and/or clinical features.
Go to :
CONCLUSION
In conclusion, this meta-analysis, which included 22,898 SLE patients and 24,838 controls, indicates that in multiple ethnic groups the functional STAT4 rs7574865 polymorphism is correlated with SLE susceptibility and that the prevalence of the rs7574865 T allele depends on ethnicity.
Go to :
 XML Download
XML Download