

 PDF
PDF Citation
Citation Print
Print



Introduction
Alzheimer’s disease (AD) is a complex, progressive, and chronic neurodegenerative condition that leads to learning, cognitive and behavioral impairment that results from neuronal and synaptic deficits in essential brain areas [1, 2]. Neuritic plaques comprising β-amyloid (Aβ) peptide as well as intraneuronal neurofibrillary tangles composed of aggregated hyperphosphorylated microtubule-associated protein tau, extracellular amyloid are parts of the pathological observations associated with AD [3]. Most studies on AD focus on the cerebral cortex and hippocampus with undoubtedly established associated roles in learning, memory, and behavioral functions, consequently causing researchers to pay less attention to the cerebellum and other areas of the brain during the clinical onset as well as the progression of AD pathology. Despite the emergence of investigative studies on the cerebellum in AD pathology suggesting that degenerative changes within the cerebellum may be inclusive in both pathogenesis and symptomatic manifestation of AD, there is still a paucity of comprehensive scientific evidence to substantiate the neuropathological observations occurring in the cerebellum of AD subjects [4, 5]. Such scientific evidence is imperative for isolating potential therapeutic agents against neuronal loss in AD and preventing resulting behavioral deficits.
Several data have described the roles of aluminum chloride (AlCl3) in the etiology of AD. Al, as an element is environmentally ubiquitous, its administration has been reported to induce neurological pathology via several routes, such as provoking dysfunction and activation of glial cells, which play an indispensable role in the regulation of the central nervous system homeostasis and neurodevelopment [6, 7]. Other routes include the generation of oxidative stress, depletion of reduced glutathione, direct and indirect reductions in mitochondrial performance and functional integrity, and increasing the production of pro-inflammatory cytokines in both the brain and peripherally [8]. Behavioral, neuroanatomical, and neurochemical alterations that mimic AD are established characteristic pathological hallmarks of both acute and chronic exposure to Al. Al promotes the formation and accumulation of insoluble Aβ and hyper-phosphorylated tau [7]. To some extent, Al creates a deficit in cortical cholinergic neurotransmission, while it increases Fe-induced oxidative injury, as seen in AD [9]. The toxicity of Al to plants, aquatic life, and humans may share common mechanisms, including disruption of the inositol phosphate system and calcium dysregulation. [10, 11].
Cymbopogon citratus, also known as lemongrass, is a tropical phytomedicinal plant that has been widely explored for its therapeutic potentials. Studies have previously reported its hypoglycemic, hypolipidemic, anxiolytic, sedative, antioxidative, and anti-inflammatory effects [12-16]. In past years, the phenolic composition of lemongrass infusion was described to include: 3-feroylquinic acid, neochlorogenic acid, chlorogenic acid, p-coumaric acid, carlinoside, isoschaftoside, isoorientin, cynaroside, veronicastroside, luteolin 7-O-neohesperidose, kurilensin A and cassiaoccidentalin B [17]. Flavonoids, such as luteolin, apigenin, glycosides, and tannins, strongly contributed to the antioxidant and anti-inflammatory properties of lemongrass [18]. The anti-inflammatory and analgesic potential of lemongrass infusion has also been reported [19].
Biological effects of flavonoids imply their bioactivity and bioavailability, and their ability to effectively reach target tissues. Therefore, using behavioral, histochemical and biochemical approaches to investigate whether extracted essential oil (ESO) of lemongrass can modulate disrupted antioxidant enzymes, altered cholinergic neurotransmission, dysfunctional neuronal bioenergetics, induction of irregular protein synthesis and neuronal cell death in cerebelli of Wistar rats were the primary objectives of the present study. Here, we have characterised the effects of chronic AlC3 exposure on the cerebellum of Wistar rats at multiple levels of organisation and complexity, while investigating the preventive and therapeutic potential of ESO in vivo.
Go to : 
Materials and Methods
Extraction of essential oil of Cymbopogon citratus
The freeze-dried sample of lemongrass (200 g; obtained from a plantation at Oke-Ose area of Ilorin in October of 2016) was steam-distilled for 1.5 hours in a Clevenger-type apparatus, with 700 ml of water at fixed temperature and extraction time based on the optimisation range to give a mixture of water/ESO. The ESO was cooled and separated from the water layer by using dichloromethane and dried in excess of anhydrous sodium sulfate. It was then stored in the dark at 4°C. Salt of AlCl3 was dissolved in distilled water to produce 20 mg/ml of AlCl3 solution and adjusted to pH 7.4.
Animal grouping and treatments
A total of 40 male rats (185±3 g) were randomly assigned into five groups (A–E, n=8) and treated orally as follows: A–phosphate-buffered saline (PBS) (1 ml daily for 15 days); B–ESO (50 mg/kg daily for 15 days); C–AlCl3 (100 mg/kg daily for 15 days); D–AlCl3 then ESO (100 mg/kg AlCl3 daily for 15 days followed by 50 mg/kg ESO daily for subsequent 15 days); E–ESO then AlCl3 (50 mg/kg ESO daily for 15 days followed by 100 mg/kg AlCl3 daily for subsequent 15 days).
Open field test
The experimental animals in this study were subjected to the open field behavioral paradigm 24 hours after the final administration. The open field apparatus was made from plywood measuring 100×100 cm with walls 50 cm high. The floor was divided into square grids, each measuring 25 cm in length with a blue marker, and a center square of the same length was drawn with a red marker. During the test, the rats were picked by their tails and dropped in the center square and allowed to explore for 5 minutes while a video was being captured by a camera from above the apparatus. Five behaviors were scored from outcomes, and these include the number of lines crossed, center square entry, center square duration, rearing frequency, and stretch-attend posture. The number of lines crossed was the frequency with which the rats crossed one of the grid lines with all four paws. The center square entry was the frequency with which the rats crossed one of the red lines with all four paws into the central square while the center square duration was the total time spent in the center square. The rearing frequency was the number of times the animal stood on its hind limbs. The stretch-attend posture is a good indicator of exploratory-anxiety conflict situations, and this was assessed by the frequency with which the animal demonstrated forward elongation of the head and shoulders followed by retraction to the original position.
Tissue processing for histological and histochemical analyses
A day (24 hours) after the last treatment, rats for histological and histochemical studies (3 from each group) were euthanised by intramuscular injection of 50 mg/kg of ketamine and subjected to transcardial perfusion during which a flush of 50 ml of normal saline was followed by 500 ml of 4% paraformaldehyde (PFA). The brain tissues were thereafter excised and post-fixed in 4% PFA for 48 hours and then processed for qualitative studies. PFA-fixed tissue sections were stained using Haematoxylin and Eosin (H&E) and Cresyl Fast Violet staining techniques. Rats processed for enzymatic studies (5 from each group) were sacrificed by cervical dislocation (to eliminate the interference of anesthetic agent with biochemical redox); the brains were then excised, rinsed in 0.25 M sucrose three times for 5 minutes each and stored in 30% sucrose at 4°C.
Biochemical assays for enzymatic analyses
Enzymatic assay for malondialdehyde (MDA), superoxide dismutase (SOD), catalase (CAT), nitric oxide (NO), glutathione peroxidase (GPx), glucose-6-phosphate dehydrogenase (G6PDH), lactate dehydrogenase (LDH), Glucokinase, acetylcholinesterase (AChE), total protein (TP) and transferrin receptor protein (TRP) were quantified in brain tissues of rats using spectrophotometric techniques at the Central Research Laboratory, University Road, Ilorin, Kwara State, Nigeria. All assay kits of the aforementioned parameters were products of Abcam, USA, except indicated otherwise. Equal weighing brain tissues were homogenised in 0.25 M sucrose at 40°C. Tissue homogenate was centrifuged for 15 minutes at 5,000 rpm, after which the supernatant containing tissue lysate was aspirated into fresh tubes. The activities of proteins were assayed according to the manufacturer’s instruction in the assay kit pack.
Photomicrography and statistical analysis
Photomicrographs of the cerebellum were obtained using an Amscope microscope camera attached to a light microscope. Quantitative analysis of the photomicrograph was done using Image J software (National Institutes of Health, Bethesda, MD, USA). The data obtained from the neurobehavioral and enzymatic assay were subjected to statistical analysis using Graph-pad Prism (version 6; Graph Pad, San Diego, CA, USA). The values were plotted in ANOVA with Tukey’s multiple comparison tests. Data obtained were presented as mean±standard error of mean (SEM) with the level of significance set at a P-value less than 0.05. The results obtained were represented in bar charts with error bars to show the mean and SEM, respectively.
Go to :
Results
ESO reversed AlCl3-induced dysregulation of general motor functions and exploratory drive in rats
The most basic and common outcome of interest in the open-field test (OFT) is movement; however, this can be influenced by motor output, exploratory drive, anxiety levels, sickness, relative time in the circadian cycle, among many other variables [21]. In Fig. 1, a similar trend can be seen in the result between the PBS and ESO groups. Rats from these groups had a significantly higher frequency in the number of lines crossed (exploration and locomotion) (Fig. 1A) and in rearing behavior (Fig. 1B) respectively than rats from the other two groups. High frequency of lines crossed and rearing is an indication of increased locomotion and exploration with a complimentarily lower level of anxiety. In addition, rats that received ESO following pretreatment with AlCl3 showed significant improvement in exploratory activities and rearing behavior (P<0.05) than rats that received AlCl3 without ESO therapy. Subsequent analysis of the frequency of central square entry and duration of time spent in the central square (Fig. 1C, D) revealed identical results between the PBS and ESO groups, indicating high exploratory habit, emotional stability and lack of anxiety in rats so treated. However, rats in the AlCl3 group exhibited a lack of verve in the open field as they merely made entry to the central square or tarried within the square (indicative of anxiety and poor motor functions), with entry frequency and duration of rats in the central square in the group significantly lower than in both PBS and ESO groups (P<0.05). Similarly, both central square entry and duration of time spent in central square of rats in the AlCl3+ESO group were lower than seen in the PBS and ESO groups. However, the frequency of central square entry and duration in the group is significantly higher than in the AlCl3 group (P<0.05). This result indicates that ESO improved exploratory behavior and lowered anxiety levels associated with AlCl3 neurotoxicity respectively. Additionally, observation of stretch-attend postures (SAP) in rats across the experimental groups revealed the significantly higher frequency of SAP in the AlCl3 group when compared with ESO and PBS groups (both at P<0.005) as shown in Fig. 1E.
 | Fig. 1(A–E) Effects of ESO administration on exploratory drive and locomotor activities in treated animals. (A) Analysis of the number of lines crossed by rats in the open field test showing no significant difference between the numbers of lines crossed by PBS and ESO controls. AlCl3 treated rats crossed the least number of lines relative to other groups. AlCl3+ESO and ESO+AlCl3 groups significantly crossed more lines than the AlCl3 treated rats (both at P<0.05). (B) Analysis of central square entry frequency in the open field test showed that rats treated AlCl3 made the least entry to the central square of the open field compared to all of PBS (P<0.05), ESO (P<0.05), AlCl3+ESO (P<0.05), and ESO+AlCl3 (P<0.05) groups. (C) Analysis of central square duration revealed that the administration of AlCl3 reduced the time rats spent in the central square when compared to the PBS (P<0.05) and ESO (P<0.05) controls. However, ESO significantly improved the central square duration of rats in the ESO+AlCl3 treated rats (P<0.05) compared to the AlCl3 group. (D) Rearing frequency analysis of experimental animals showing significant reductions in both groups treated AlCl3 compared to other groups. ESO intervention improved rearing frequencies in rats in the AlCl3+ESO and ESO+AlCl3 groups (at P<0.05, respectively) compared to the AlCl3 group. (E) Analysis of the stretch-attend posture frequency shows significant elevation in rats treated AlCl3 treated compared to PBS and ESO control (both at P<0.05). However, the AlCl3+ESO and ESO+AlCl3 treated rats showed improved stretch-attend-posture, which is significantly different (both at P<0.05) from the AlCl3 group. The results are presented as mean±standard error of mean. AlCl3, aluminum chloride; ESO, essential oil; PBS, phosphate-buffered saline. *P<0.05.
|
Effects of ESO on AlCl3-induced perturbation of cerebellar redox
Aluminum is known to cause neurotoxicity by exacerbating the production of reactive oxygen and nitrogen species. In this study, the AlCl3 group presented with significantly depleted level of SOD when compared to the control group (P<0.05). Interestingly, the aluminum group presented with a raised level of CAT and GPx. This finding suggests that AlCl3 mediated a cascade of chemical event that rsulted in the elevated levels of superoxides, which led to a depletion in the level of SOD. To further examine the beneficial potentials of ESO in reversing AlCl3-induced oxidative stress, we measured cerebellar lysate levels of CAT, GPx, and SOD in the ESO pre and post-teated groups (Fig. 2). The results revealed that ESO significantly improved SOD activities in the AlCl3+ESO (P<0.05) and ESO+AlCl3 (P<0.05) when compared to the AlCl3 group.
 | Fig. 2Cerebellar oxidative redox state characterized by (A) SOD, (B) CAT, (C) GPx, (D) MDA, and (E) NO activities respectively. (A) Cerebellar SOD activity was significantly depleted by AlCl3 treatment relative to both PBS (P<0.05) and ESO groups (P<0.05) but was normalized by ESO treatment in ESO+AlCl3 (P<0.05) and AlCl3 (P<0.05) groups. (B) CAT level was increased by AlCl3 relative to both PBS and ESO controls (P<0.05) but regularized by ESO treatment in both AlCl3+ESO and ESO+AlCl3 groups at P<0.05. (C) AlCl3 treatment induced an increase in GPx level relative to the PBS and ESO groups at P<0.05 for both. ESO treatment restored the level of GPx in AlCl3+ESO (P<0.05) to normal levels but not ESO+AlCl3 group. (D) AlCl3 treatment triggered a significant increase in the level of MDA when compared to the PBS (P<0.05) and ESO (P<0.05) controls. This was normalized in the AlCl3+ESO (P<0.05) and ESO+AlCl3 (P<0.05) groups. (C) AlCl3 treatment elicited a striking increase in NO activities relative to the PBS and ESO groups (both at P<0.05) and was regularized by ESO in both AlCl3+ESO and ESO+AlCl3 groups (at P<0.05 each). AlCl3, aluminum chloride; CAT, catalase; ESO, essential oil; GPx, glutathione peroxidase; MDA, malondialdehyde; NO, nitric oxide; PBS, phosphate-buffered saline; SOD, superoxide dismutase. *P<0.05.
|
Furthermore, our findings revealed that AlCl3-treatment induced lipid peroxidation and nitrosative stress in the cerebellum of rats when compared to animals administered with PBS alone, as evidenced by elevated MDA (P<0.05) and NO (P<0.05) levels in cerebellar lysates. Significant decreases in levels of MDA and NO were recorded in groups pre- and post- exposed to AlCl3 subsequent to ESO administration, both at P<0.05 for MDA and NO (Fig. 2).
Cerebellar G6PDH, LDH, and glucokinase expressions following AlCl3 and ESO treatments in rats
Tissue levels of G6PDH were significantly reduced by AlCl3 treatment in the cerebellum (P<0.005) relative to the PBS and ESO controls. Both pre- and post-administrations of ESOs were able to effectively counteract the neurotoxicity of AlCl3 by regulating cerebellar G6PDH to baseline levels (Fig. 3A). To further ascertain the protective effect of ESO, we measured LDH expression within cerebellar lysates under different conditions. AlCl3 oral infusion significantly increased LDH activity within the cerebellum, which is an index of neurotoxicity (Fig. 3B). We found ESO to be significantly effective in normalizing LDH expression to baseline levels in both AlCl3+ESO (P<0.05) and in the ESO+AlCl3 (P<0.05) groups. Further analysis showed that AlCl3 treatment depleted the level of Glucokinase in cerebellar lysates, although changes were insignificant statistically (Fig. 4). ESO treatment was, however, able to restore the level of Glucokinase in the cerebellum back to basal levels, i.e. as compared with the control (PBS and ESO).
 | Fig. 3Cerebellar G6PDH (A), LDH (B), GSA (C), TRP (D), and AChE (E) activities under treatment conditions. (A) AlCl3 treatment caused depletion in G6PDH expression relative to the PBS and ESO groups (P>0.05 in both). Pre- and post-treatment with ESO normalized G6PDH levels in the AlCl3+ESO (P<0.05) and ESO+AlCl3 (P<0.05) groups. (B) Expectantly, AlCl3 treatment induced a significant decrease in the level of LDH compared to the PBS, and ESO controls both at P<0.05. ESO treatment prevented LDH dysregulation in AlCl3+ESO (P<0.05) and ESO+AlCl3 (P<0.05) treated rats. (C) Cerebral GSA activity was significantly depleted in AlCl3 treated rats when compared to PBS (P<0.05), ESO (P<0.05), AlCl3+ESO (P<0.05) and ESO+AlCl3 (P<0.05) treated rats. (D) The expressed level of TRP in AlCl3 treated rats was significantly higher than in PBS, and ESO controls at P<0.05 each, as well as in AlCl3+ESO and ESO+AlCl3 treated rats (both at P<0.05). When compared to all other treated groups, AlCl3 treated rats presented with the highest level of AChE, with significant modulation seen following ESO intervention in the AlCl3+ESO and ESO+AlCl3 both at P<0.05. AChE, acetylcholinesterase; AlCl3, aluminum chloride; ESO, essential oil; GSA, glucokinase specific activity; G6PDH, glucose-6-phosphate dehydrogenase; LDH, lactate dehydrogenase; PBS, phosphate-buffered saline; TRP, transferrin receptor protein.*P<0.05.
|
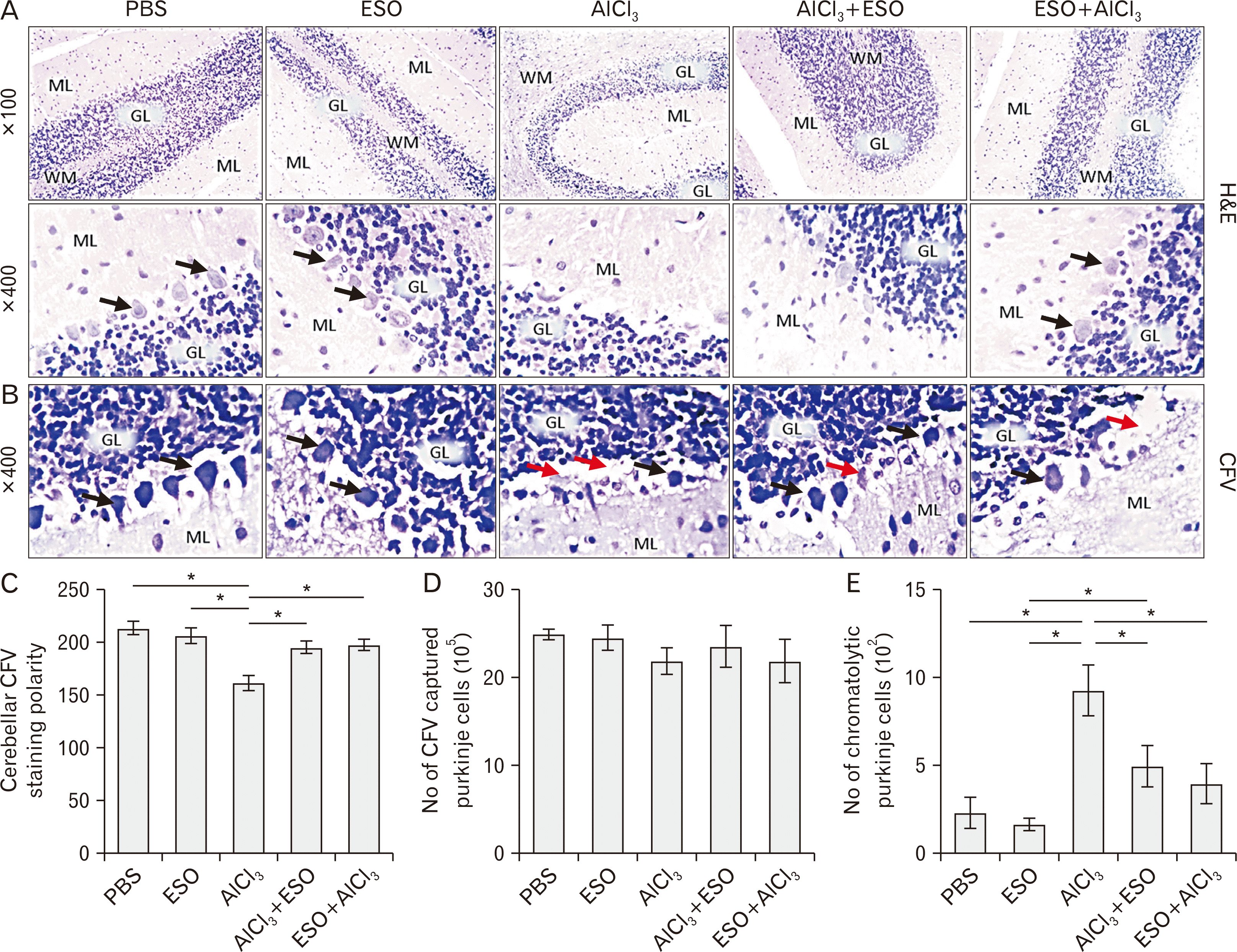 | Fig. 4(A–E) Representative photomicrographs showing panoramic views of cerebellar cortex general morphological presentations and Nissl profile in Wistar rats across the various study groups. (A) is H&E (×100, ×400); The ML, Purkinje cells (black arrowhead) layer, GL, and the medullary layer of WM are demonstrated across study groups. PBS and ESO treatments did not alter the morphological presentation of cerebellar layers from this study. Both groups show a fine array of cells that are distinctly arranged from the molecular to the granular layer. Also, cellular density within the groups appears normal across all cortical layers. AlCl3 treatment caused degenerative changes characterized by fragmented GL in cerebellar cortex and poorly stained Purkinje cells. Comparatively, increased cellular density was observed in this group. Groups administered ESO before and after AlCl3-induced toxicity (AlCl3+ESO and ESO+AlCl3) shows mild degenerative changes in line of morphologic appearance of cerebellar sections of AlCl3-treated rats; cerebellar layers in both groups looks better structured and layers are well-delineated. (B) shows Nissl profile as demonstrated by CFV stain (×400). Normal morphological presentations of the cerebellar cortex are observable in PBS and ESO treated groups that are characterized by well-defined staining profiles. Cellular morphology in these groups is characterized by Purkinje cells with conspicuous cell bodies and dendrites that are projecting deep into the MLs (black arrowhead). Also, the granule layer in these groups consists of small granule neurons which are well outlined in contrast to the loosely arranged and cryptic cells in the granule layers of groups AlCl3 treated groups. Degenerating Purkinje cells with chromatolytic cell bodies (red arrowhead) and short dendritic processes can be seen around the indistinctly demarcated cerebellar layers of groups that receive aluminum treatment. The transitional regions between the cerebellar layers of AlCl3 treated with ESO are also generally better delineated. Although a few perineural spaces are present around Purkinje neurons of this groups, the general cytoarchitecture is much similar to that of the PBS and ESO groups. (C–E) show quantitative analysis of the CFV stain. AlCl3, aluminum chloride; CFV, Cresyl Fast Violet; ESO, essential oil; GL, granule cell layer; ML, molecular layer; PBS, phosphate-buffered saline; WM, white matter.*P<0.05.
|
ESO Modulates AChE and TRP activities following AlCl3-induced neurotoxic cascades
The loss of cholinergic neurotransmission in the cerebral cortex and other areas contribute significantly to deterioration of cognitive function seen in patients with AD. AChE interferes with choline uptake/release at presynaptic terminals, resulting in an alteration of neurotransmission evidenced by substantial neocortical deficits in the enzyme responsible for the synthesis of acetylcholine (ACh), choline acetyltransferase (ChAT). We observed elevated levels of AChE in AlCl3-treated groups (Fig. 4A) relative to both PBS and ESO controls. Similar to earlier results of this study, we found that pre- and post- treatment of rats with ESO following AlCl3-induced neurotoxicity significantly downregulated AChE levels to basal levels (both at P<0.01). In rats that received AlCl3, we further observed a marked increase in the level of TRP (Fig. 5C) when compared to baseline levels in the controls (PBS and ESO). The therapeutic potentials of ESO were further buttressed in this study by its normoregulating roles in expressed TRP in cerebellar homogenates, wherein animals post-treated with ESO following AlCl3 infusion (AlCl3+ESO) showed significantly decreased TRP expression in comparison to the AlCl3 (P<0.05 and P<0.01 respectively).
Cerebellar histology and nissl profile following AlCl3-induce toxicity and d ESO therapy
The cerebellum possesses distinctive histological appearance that can become invariably altered secondary to inflammatory reactions and oxidative stress dysregulation. We comparatively examined thin sections of cerebellar tissues using routine H&E and Nissl staining methods. Panoramic representation of cerebellar histological profile of the control groups (PBS and ESO) was regular, with consistent layering and succinct transitions seen between molecular, granular and Purkinje neuron layers (Fig. 4A). Similarly, a close examination of the Purkinje cells using a higher magnification, showed that neither of PBS nor ESO treatment altered the structural integrity of these all-important cerebellar neurons. Additionally, apical and basal dendrites of neurons were well expressed and interconnected within the neuropil of both control groups (Fig. 5A). As anticipated, the cerebellar milieu of AlCl3-treated rats were characterised by the extravasation of the granule cells into the Purkinje cell layer, fragmentation of neuropil, distortion of layering, and cytoplasmic degeneration of neurons. Furthermore, there was a marked reduction in the deposition of Nissl proteins in cerebellar sections of these groups when compared to rats that received PBS and ESO, which is indicative of chromatolytic changes via alteration in protein synthesis (Fig. 4B). Quantitative analysis of the Nissl stain revealed that the aluminium group had a significantly reduced staining intensity when compared to the control group (P<0.05) (Fig. 4C). Also, relative to the control group, the AlCl3 group presented with more chromatolytic Purkinje neurons (Fig. 4E). Interestingly, both treatments with ESO before and after AlCl3-induced degenerative changes on the cerebellum improved general histomorphology and Nissl profiles, as seen in both AlCl3+ESO and ESO+AlCl3 groups (Fig. 4B). The AlCl3+ESO and ESO+AlCl3 groups also presented with an increase in staining intensity and reduced Purkinje chromatolysis when compared to the AlCl3 group. The finding suggests that ESO can restore/maintain histomorphological integrity to the cerebellum following AlCl3 induced neurotoxicity.
Go to :
Discussion
This study showed the possible neuroprotective/neuromodulatory potentials of ESO before and after AlCl3 exposure on rat’s cerebellar morphology and functions through behavioral, biochemical, and histoarchitectural evaluations. The use of neurobehavioral studies in risk assessment is premised on the fact that behavior/emotions can be regarded as the net output of memory, motor, locomotor and cognitive functions occurring in the nervous system and can serve as potentially sensitive endpoints of chemically induced-neurotoxicity [22]. AlCl3 is a neurotoxicant that permeates and functionally alters the blood-brain barrier and potentially produces changes in the ionic, cholinergic, dopaminergic, and noradrenergic neurotransmission in the central nervous system. These alterations are known to be associated with impairment of visuoperception, attention, cognitive, memory, and learning ability [23, 24]. Data obtained from the OFT showed a decrease in exploratory activities of rats treated with AlCl3, as seen by a significant decline in the number of lines crossed, rearing frequency, center square entry, center square duration, and elevated SAP when compared with other groups. Reports of decreased exploratory and locomotor activities have been reported following AlCl3 treatment. Our lab and others from literature have reported decreased crossing scores and altered secondary parameters such as rearing, grooming, and sniffing in the OFT following AlCl3 treatment [25, 26]. Decreased locomotor activity assessed by actophotometer following treatment with AlCl3 was reported, and it was deduced from this study that this effect might be due to possible depressant effects of AlCl3 on the CNS [23]. A significant reduction has been documented in the exploratory habits of rats treated with sodium azide–another mitochondrial toxin with neurotoxicant properties [27]. Suggestively, the altered behavioral pattern seen in the AlCl3 group in this study might be due to its ability to impair glucose utilisation and the release of cytochrome C, which inhibits the production of ATP generation. This hypothesis is supported by findings of glucose bioenergetics dysfunction seemingly caused by AlCl3 treatment in this study. ESO treatment before and after AlCl3 toxicity significantly improved exploratory and locomotor outcomes in the OFT. It is noteworthy that treatment with ESO before AlCl3 treatment was more effective than after, indicating that ESO is more effective in a preventive/pro-adaptive/protective role. Improved glucose biosynthesis parameters in the cerebellum of rats treated with ESO following AlCl3 toxicity may underlie its therapeutic and protective efficacy against motor-related behavioral alterations in this study.
The mechanism of cytotoxic and/or neurotoxic substances are thought to be mainly due to the oxidative stress involved in the production of reactive oxygen species (ROS), including superoxide anion, hydrogen peroxide, superoxide radical and hydroxyl radical. Moreover, the degree of oxidative damage is depended on the balance between the oxidative stress and the efficiency of the endogenous antioxidant system that is found in the majority of cells [5]. Generally, brain tissues are highly susceptible to attacks from free radicals due to highly unsaturated lipid contents [28]. Studies have shown that exposure to Al results in the impairment of mitochondrial functions in vivo and in vitro, as well as destroying the antioxidant defense system by decreasing the antioxidant enzyme status [29]. We, therefore, measured lipid peroxidation by assessing MDA levels, nitrosative stress by quantifying NO and determining status of endogenous antioxidant enzymes, including SOD, CAT, and GPx following AlCl3-induced cerebellar toxicity while studying the possible neuroprotective mechanisms of ESO. Since lipid oxidation products are one of the primary outcome associated with oxidative stress, the significantly increased cerebellar level of MDA in the group treated with AlCl3 in this study reflect induction of lipid production by the neurotoxicant. This result corresponds with other studies in which Al treatment increased iron-dependent lipid peroxidation in rat brain [30], and especially in cerebrum and cerebellum following intraperitoneal treatment of Al [26]. In this study, rats treated with ESO before and after AlCl3 intoxication show marked reduction of MDA level in the cerebellar homogenate. Cymbopogon citratus has been reported to possess flavonoids, such as luteolin, apigenin, glycosides and tannins and these actively contributed to its antioxidant and anti-inflammatory properties [18], and this may underlie its ability to reduce lipid peroxidation in this study.
Overproduction of NO makes it noxious and can undergo oxidative–reductive reactions if a cell is in a pro-oxidant state to form toxic compounds (reactive nitrogen species’, or RNS), which cause cellular damage [31, 32]. Recently, the term ‘nitrosative stress’ has been used to indicate the cellular damage that is elicited by excess NO and RNS (peroxynitrite and nitrogen [III] oxide), and have been implicated in the pathogenesis of neurodegenerative disorders. Nitrotyrosine formation, a marker of nitrosative stress, has been reported both in AD and Parkinson’s patients [33]. As a response to AlCl3-induced neurotoxicity in this study, we observed significant upregulation of NO in the cerebellar cortex of rats, which might be due to the ability of Al to selectively favor the production of NO from l-arginine via the activation of NO synthase. Interestingly, we found that rats treated with ESO before and after AlCl3 intoxication shows normal NO level in cerebellar lysates, which correlates with improved behavioural outcome.
This observation was further buttressed by results from SOD, CAT, and GPx analyses. Recent evidence suggests the formation of superoxide anion and expression/activity of its endogenous scavenger-SOD as a common player in the pathogenic cascades of many neurodegenerative pathologies like AD [5]. Similarly, GPx an enzyme that reduces lipid hydroperoxides to their corresponding alcohols and reduces free hydrogen peroxide to water, and CAT, which protects SOD by converting H2O2 to water and oxygen, have both been implicated in AD etiology. Our findings showed that treatment with Al evidently increases oxidative stress and brain injury as there was significant repression in the levels of SOD, GPx, and CAT in the cerebellar homogenate relative to control. These results corroborated previous findings that Al treatment results to downregulation of antioxidant enzymes, causing an imbalance between pro-oxidant and antioxidant potentials [5, 26]. On the contrary, the administration of ESO before and after Al treatment showed a reversal of the deleterious effects on oxidative redox within the cerebellum of rats. Flavonoids are known to exert their protective effect by directly scavenging ROS, by activating antioxidant enzymes or through metal chelating activity. These may account for the relative restoration of homeostasis shown by ESO in this study.
Disturbances in central or peripheral glucose bioenergetics, which can be affected by almost every level of cellular or biochemical metabolic cascades have been implicated in various neurodegenerative pathologies [5]. LDH and G6PDH are enzymes in the energy-producing glycolytic pathway, which may be affected by oxidative modification and decreased activity may contribute to the alteration in glucose metabolism noted in AD [34]. The consequence of loss of neuronal membrane integrity due to oxidative injury is the release of LDH in a large quantity. Its aberrant production in cells is a useful tool for determining cytotoxicity and in measuring early cellular damage or impairment [35]. The cerebellar cortex of rats treated AlCl3 showed significant upregulation of LDH activities in the cerebellar cortex when compared to the control group. Reports of LDH alteration in the cerebellar cortex following treatment of rat with Al have been documented, more than any metabolically active organ, LDH gene expression ratio in the brain is the most altered in response to impaired oxidative phosphorylation [36]. Alteration of cerebellar LDH levels in this study may also be linked to oxidative stress caused by excess production of ROS and RNS induced by Al treatment. Rats in this group also experienced a decline in energy and ribose production through pentose phosphate pathways, as evidenced by decreased G6PDH in cerebellar homogenates. These findings indicate that one mechanism through which Al induces its toxicity is by altering energy production through inhibition of the activities of key enzymes responsible for glucose metabolism and that ESO has both protective and therapeutic effects against dysfunction of cerebellar bioenergetics. Transferrin is known primarily for its role in the transport and cellular uptake of iron but is also the major serum binding protein for aluminum [37]. One of the mechanisms through which Al exerts its neurotoxicity is its ability to hinder iron transport and absorption by displacing iron from binding to transferrin and therefore inhibit iron metabolism [38]. We investigated cerebellar levels of TRP following the oral infusion of Al and ESO in Wistar rats. AlCl3 upregulated TRP in the cerebellar of rats, which may be due to the ability of Al to prevent the binding of iron to transferrin resulting into the accumulation of TRP and subsequently iron-induced oxidative stress. The potentials shown by ESO in normalising aberrant TRP expression is an underpinning of its therapeutic mechanism in this study. ESO’s ability to prevent the accumulation of transferrin maybe by inhibiting Al from displacing iron in the iron-transferrin complex and subsequent iron-induced oxidative stress, which is crucial to inhibiting eventual neurotoxic cascades.
AChE plays a crucial role in learning, memory, and cognitive functions [23]. Cholinergic neurons are positive markers for the evolution of memory and related disorders affecting ACh, and impaired cholinergic transmission is evident in various neurological diseases associated with cognitive impairment [24, 26]. In addition to findings of the present study, cholinotoxic effects exerted by AlCl3 has been shown in previous studies to produce a functional change in cholinergic and noradrenergic neurotransmission. Such toxic changes to the neurotransmission system in the cerebellum may account for the loss of locomotor activities that were recorded in the open field, as we have found. Also, the ability of ESO to mitigate AChE upregulation in the cerebellum is an important aspect of its ability to restore and prevent locomotor irregularities. An interplay between increased tissue ROS and alteration of AChE synthesis has been described [23], spotlighting a possible bidirectional mechanism of action for ESO in the present study.
Histological observations showed that the cerebellar cortex in rats treated AlCl3 is characterised by fragmented granule cell layers with distorted neuropil, pyknotic Purkinje cells, and increased cellular density. Therefore, inhibition of the demonstrated cytotoxic impact of AlCl3 by ESO treatment is also a key component of its therapeutic potentials. Additionally, Nissl integrity in granule and Purkinje neurons within the cerebellum of AlCl3 treated rats show highly chromatolytic changes compared to controls, which indicates abnormal normal endoplasmic functioning and intercellular protein synthesis - corroborating results obtained from TP biochemical analysis.
Nissl granules are known to change under physiological and pathological conditions where they may dissolve and disappear in a condition known as chromatolysis, which can be triggered by axotomy, ischemia, and toxicity to the cell, as well as cell exhaustion or virus infections leading to disintegration [39]. If not checked, chromatolysis often resuly in cell death and ultimately neurodegeneration. Our present finding is in agreement with previous reports that highlighted the neurodegenerative impact of Al on the Nissl bodies in neurons [40, 41]. Al-induced trauma to protein macromolecules and induced endoplasmic reticular (ER) stress in neurons may have resulted in chromatolysis and progressive decline in normal cerebellar histomorphology and function. The observed chromatolytic changes in the Purkinje neurons may explain the behavioral decline of the animals in the OFT. Therefore, the role of ESO to prevent and reverse chromatolysis in cerebellar neurons in this study may stem from its ability to prevent ER stress and inhibiting pathways that lead to aberrant protein production owing to its antioxidative and ROS scavenging properties.
In conclusion, summarily, our results strengthen the idea that AlCl3 initiates neuronal toxicity and degeneration in the cerebellum through the initiation of oxidative stress, as seen by the excessive production of free radicals and depleted endogenous antioxidant system, which resulted in neuronal damage. It is, however, intriguing that these degenerative changes were pharmacologically mitigated following ESO administration and should, therefore, be explored further towards the development of novel therapy for neurodegeneration.
Go to :
 XML Download
XML Download