

 PDF
PDF Citation
Citation Print
Print



Introduction
Obesity, a state of excess accumulation of white adipose tissues (WATs), and its associated metabolic diseases are increasing worldwide. Adipose tissues play important roles in systemic metabolism by storing and releasing energy as well as acting as endocrine organ. Adipose tissues become dysfunctional in obesity, which is characterized by hypertrophied adipocytes, elevated inflammation, hypoxia and fibrosis and reduced angiogenesis [1]. Alterations in adipose derived factors, elevated levels of fatty acids (FAs) and proinflammatory cytokines along with low level of adiponectin from higher mass of dysfunctional adipose tissues, are thought to cause or exacerbate cardiometabolic diseases in obesity.
Adipose tissues are present in multiple locations throughout an organism and largely divided into intraabdominal visceral and subcutaneous depots. The major visceral depots are omental and epiploic fat and major subcutaneous depots are the upper-body abdominal subcutaneous and the lower-body femoral and gluteal fat in humans [2]. In addition to WATs, bioenergetically more active brown adipose tissue (BAT) as well as inducible brown-like adipocytes, beige or brite adipocytes, exist. Brown and beige/brite adipocytes contain more mitochondria and express uncoupling protein 1 (UCP1) and hence, have higher thermogenic capacity [3]. Each adipose depot has differential influence on systemic metabolism. Visceral adiposity, over and above total fat mass per se, is independently associated with metabolic complications, whereas fat accumulation in the lower-body is protective [2]. The amount of BAT or beige/brite fat is reduced in obesity and increasing their amount and activity could confer protection against obesity and its associated metabolic diseases [4].
In addition to its well-known effects on calcium homeostasis and bone metabolism, vitamin D exerts many other actions including proliferation and differentiation of cells and immunomodulatory functions [5]. In adipose tissues, vitamin D has been shown to affect adipocyte development and their metabolic and endocrine functions [6]. While obesity is associated with low vitamin D status and metabolic diseases, whether deficiency of vitamin D predisposes to obesity or vitamin D supplementation improves obesity and metabolic diseases is not clear. We will review vitamin D regulation of adipose biology including adipose tissue development and metabolic and endocrine properties with emphasis on molecular mechanisms that link low vitamin D status with obesity and metabolic diseases. We also discuss the effects of supplementation or maintenance of vitamin D on obesity and metabolic diseases.
Go to : 
VITAMIN D METABOLISM AND BIOLOGICAL FUNCTION
Vitamin D metabolism
Vitamin D is synthesized from 7-dehydrocholesterol in the skin (vitamin D3) or ingested as food (both vitamin D2 and D3 forms). Ultraviolet B photons cause the photolysis of 7-dehydrocholesterol to previtamin D3, which thermally isomerizes to vitamin D3. Vitamin D (D represents D2 and/or D3) is activated through 2 hydroxylation steps (Fig. 1). In the liver, vitamin D is hydroxylated to 25-hydroxyvitamin D [25(OH)D] by 25-hydroxylases (CYP2R1, CYP27A1, CYP3A4, CYP2J2) [7]. 25(OH)D is then activated to 1,25 dihydroxyvitamin D [1,25(OH)2D] by 1α-hydroxylase (CYP27B1) in the kidneys. Vitamin D binding protein is the specific chaperone for vitamin D and its metabolites in the blood and then to the storage sites (adipose tissues and skeletal muscle) or target tissues (liver, kidneys or parathyroid gland) and cells (monocytes and macrophages). Both 25(OH)D and 1,25(OH)2D are hydroxylated by 24-hyroxylase (CYP24A1) and degraded. Other tissues including adipose tissues also express 1α-hydroxylase and 25-hydroxylase and can activate vitamin D locally [8].
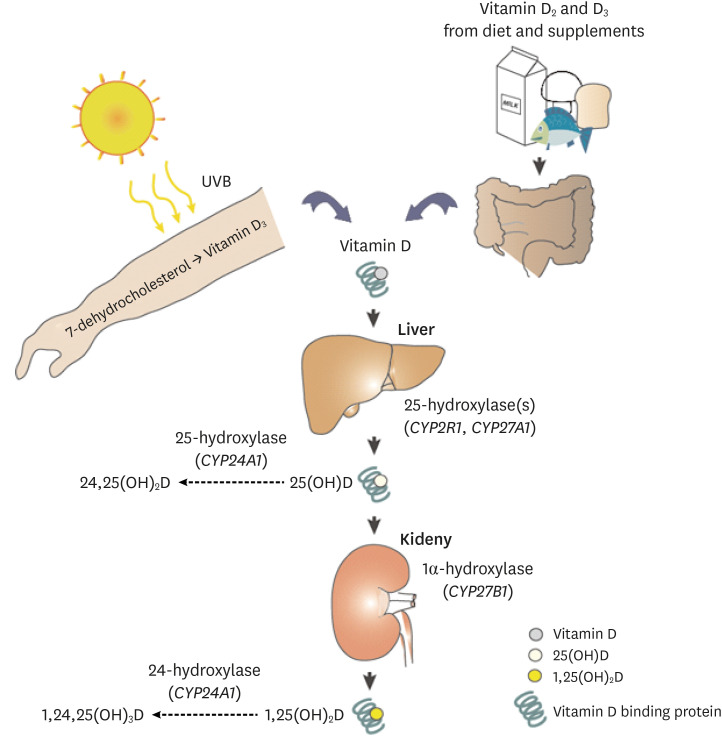 | Fig. 1Metabolism of vitamin D. Vitamin D, synthesized in the skin from 7-dehydrocholesterol as D3 or ingested from food in the forms of D2 or D3, are first converted to 25(OH)D (D2 or D3) by 25-hydroxylases (encoded by CYP2R1, CYP27A1, CYP3A4, CYP2J2) in the liver. 1α-hydroxylase (CYP27B1) converts and activates 25(OH)D to 1,25(OH)2D in the kidneys. Other tissues and cells express 1α-hydroxylase and can activate 25(OH)D to 1,25(OH)2D. Vitamin D and its metabolites are bound to DBP in the blood. Both 25(OH)D and 1,25(OH)2D are catabolized by 24-hydroxylase (CYP24A1).UVB, ultraviolet B; DBP, vitamin D binding protein.
|
Biological functions of vitamin D
In addition to regulation of calcium homeostasis and bone metabolism, vitamin D regulates broad biological processes including proliferation, differentiation and maturation of cells, immune functions and cellular metabolism. We refer to other reviews for the roles of vitamin D and we will briefly describe the molecular details through which vitamin D regulates cellular actions.
Most of biological actions of 1,25(OH)2D3 are thought to be mediated through its nuclear receptor, vitamin D receptor (VDR) (Fig. 2). Ligand bound VDR translocates into the nucleus as a heterocomplex with retinoid X acid receptor and controls gene transcription by binding to vitamin D response elements of genes [9]. Through interactions with other nuclear receptors including nuclear factor kappa B (NF-κB), SP1 and STAT5 vitamin D controls transcription of genes. 1,25(OH)2D3-VDR regulates hundreds of genes in cell- and tissue-specific manners [10].
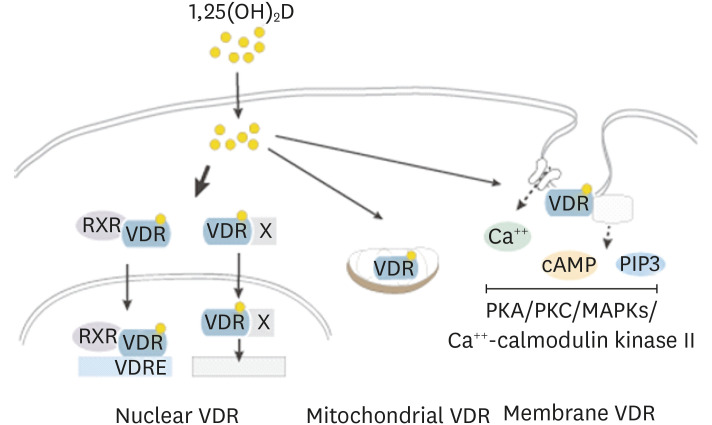 | Fig. 2Vitamin D exerts biological functions through the VDR. The active form of vitamin D, 1,25(OH)2D, is a relatively small, lipophilic molecule that easily penetrates the cell membrane by simple diffusion and binds to VDR. Upon binding to the ligand, VDR heterodimerizes with RXR and translocates into the nucleus, where it controls gene transcription by binding to VDREs. VDR interacts with other nuclear receptors (X) and regulates transcription of genes. VDR is present in the cavaeoli of plasma membrane, where it exerts rapid nongenomic actions through multiple signaling pathways including PKA and PKC, MAPKs, and Ca++-calmodulin kinase II. VDR also has been shown to be present in mitochondria where it affects mitochondrial respiration.VDR, vitamin D receptor; RXR, retinoic X acid receptor; VDRE, vitamin D response element; PKA, protein kinase A; PKC, protein kinase C; MAPK, mitogen activated protein kinase; cAMP, cyclic adenosine monophosphate.
|
VDR localizes in the caveolae structures of plasma membrane where it exerts rapid membrane-initiated signaling responses that are referred as non-genomic actions of vitamin D [11] (Fig. 2). Once bound to vitamin D, VDR interacts with plasma membrane proteins including phospholipase A2, phosphatidylinositol-3 kinase and calcium transporters. These lead to generation of secondary messengers, Ca++, cyclic adenosine monophosphate, and phosphatidylinositol 3,4,5 triphosphate (PIP3) and activation of downstream protein kinase A, protein kinase C, mitogen activated protein kinases (MAPKs) and Ca++-calmodulin kinase II [11]. Through these signaling events, vitamin D is known to affect many cellular responses. These signaling events can also lead to control of gene expression through modulation of transcriptional machinery.
VDR is present in mitochondria where 1,25(OH)2D3 is known to negatively affect respiratory capacity in multiple cell types including platelets and keratinocytes [1213]. Vitamin D can also inhibit cell respiration through the nuclear VDR-mediated suppression of transcription of mitochondrial respiratory chain complexes. Through regulation of mitochondrial respiratory capacity, 1,25(OH)2D3-VDR may affect cell growth, proliferation and differentiation as well as biosynthetic pathways, especially lipid biosynthesis, as it provides bioenergetics required for the processes [13].
Vitamin D metabolism in adipose tissues
Vitamin D accumulates in adipose tissues and skeletal muscle and adipose tissue is thought to be the major site of vitamin D storage [14]. Slow releases from adipose tissues, however, suggest that adipose tissues may function as a vitamin D buffering system preventing uncontrolled synthesis of the active form. Vitamin D metabolizing enzymes, 25-hydroxylase (CYP2R1, CYP27A1, CYP2J2), 1α-hydroxylase (CYP27B1) and catabolic 24-hydroxylase (CYP24A1), are expressed in adipose tissues [15161718], indicating that adipose tissues also play active roles in vitamin D metabolism contributing to the low vitamin D status in obesity.
Expression levels of 25-hydroxylase (CYP2J2) and 1α-hydroxylase (CYP27B1) are reduced in obese adipose tissues [16], whereas VDR expression is increased in obesity and is positively associated with proinflammatory cytokine expression [161718]. Visceral than subcutaneous adipose tissues express higher levels of CYP27A1 and lower levels of CYP27B1 and CPY2J2 [16]. Furthermore, 1,25(OH)2D3 increases VDR expression in visceral adipose tissues from obese but not lean subjects [17]. These data indicate that local activation of vitamin D as well as their actions in adipose tissues may differ depending on the degree of obesity and adipose depots, potentially explaining the contradictory effects of vitamin supplementation on obesity and metabolic diseases.
Go to :
VITAMIN D AND ADIPOSE BIOLOGY
Vitamin D regulation of adipogenesis
Adipose tissues continuously remodel throughout a life-span and mean age of adipocytes is estimated to be about 10 years in humans [19]. Adipose tissues remodel by increasing the size (hypertrophy) and or number (hyperplasia) of adipocytes. Remodeling through hyperplasia is considered to be protective as newly-differentiated adipocytes are insulin sensitive and contain higher capacity to store excess energy, protecting other organs from ectopic fat deposition [20]. In contrast, hypertrophic obesity is associated with impairment in adipogenesis, high inflammation and fibrosis, markers of dysfunctional adipose tissues [2122]. Replacing dysfunctional adipocytes during development of obesity and normal aging through new adipocyte generation is crucial for the maintenance of metabolic health. We will review current understanding of the role of vitamin D in adipogenesis.
Controversial effects of vitamin D on adipogenesis
Adipogenesis is a process through which adipose progenitors differentiate into mature adipocytes. Upon treatment with adipogenic stimuli, adipose progenitors start to express adipocyte specific genes and significant changes in morphologies occur such that cells round up and accumulate neutral lipids in the form of triacylglycerol (TAG) in lipid droplets [23]. To assess the direct effects of vitamin D on adipogenesis, the active form of vitamin D, 1,25(OH)2D3, has been added to various cell culture models including 3T3-L1, a mouse embryonic cell line, and primary cultures of adipose derived stem cells (ASCs). Results from these in vitro studies, however, are controversial depending on the experimental conditions, cell types or doses used (Fig. 3). In 3T3-L1 cells, 1,25(OH)2D3 inhibits adipogenesis [2425] while enhancing adipocyte differentiation in human and mouse ASCs [152627] and in bone marrow-derived mesenchymal stem cells (BM-MSC) from mouse [26] and pigs [28]. Differences in adipogenic programs between cell types may explain the contradictor results [23].
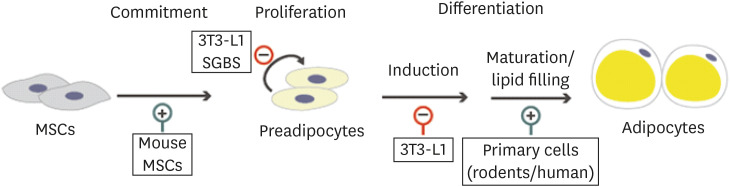 | Fig. 3Vitamin D regulation of adipogenesis. Effects of vitamin D on each step of adipogenesis in different cell types are summarized (⊕, stimulatory effect; ⊖, inhibitory effect). Vitamin D directs MSCs to adipogenic lineage through the vitamin D receptor-dependent mechanisms. Vitamin D has been shown to suppress proliferation in both 3T3-L1 and SGBS cells, dose-dependently. In 3T3-L1 cells, vitamin D inhibits adipogenesis by acting on early states of differentiation. In contrasts, vitamin D enhances differentiation of primary preadipocytes isolated from human and rodent adipose tissues by acting on later maturation steps.MSC, mesenchymal stem cell; SGBS, Simpson-Golabi-Behemel Syndrome.
|
Preadipocytes are known to express vitamin D metabolizing enzymes and locally activated 1,25(OH)2D3 may also modulate adipogenesis. We demonstrated that 25(OH)D3 increases the expression levels of CYP24A1, a primary target of nuclear VDR, and enhances adipogenesis in human ASCs, indicating that they can generate the active form of vitamin D from 25(OH)D3 [15]. Consistent with this idea, we detected 1,25(OH)2D3 in the culture media when human ASCs are incubated with 25(OH)D3 [15]. Vitamin D did not increase CYP24A1 expression, suggesting that 25-hyroxyalse may not be functional in human ASCs.
Molecular mechanisms through which 25(OH)D3 regulates adipogenesis
Both pro-adipogenic and anti-adipogenic actions of vitamin D are known to be mediated through the VDR. Kong et al. [25] showed that 1,25(OH)2D3 inhibits adipogenesis through the VDR-dependent suppression of PPARγ, the master regulator of adipogenesis, in 3T3-L1 cells. In contrasts, 1,25(OH)2D3 stimulates adipogenesis in BM-MSC derived from the wild-type, but not from VDR-knockout mice and introduction of human VDR into the knockout cells rescued the pro-adipogenic effects of 1,25(OH)2D3 [26]. Whether 1,25(OH)2D3 promotes adipogenesis in human and mouse ASCs through VDR dependent mechanisms has not been demonstrated.
Vitamin D is known to affect cell proliferation dose-dependently in both 3T3-L1 and Simpson-Golabi-Behemel Syndrome (SGBS) cells, a primary human preadipocytes obtained from subjects with SGBS [2729]. By inhibiting cell proliferation, vitamin D may affect adipogenesis. Consistent with this idea, 1,25(OH)2D3 is inhibitory when added during the earlier days of differentiation but does not affect when added during later days of adipogenesis in 3T3-L1 cells [25], a cell line that post-confluent mitotic clonal expansion is critical for their differentiation into adipocytes. On the contrary, we and others have shown that 1,25(OH)2D3 increases lipid accumulation by acting on the later periods of adipogenesis without affecting cell commitment in human and mouse ASCs [1530]. 1,25(OH)2D3 may support the differentiated state after induction of adipogenesis is consolidated by stimulating FA biosynthesis and lipid accumulation as acetyl-CoAs are directed for synthetic pathway from oxidation in mitochondria [13].
Lessons learned from mouse models
To understand roles of vitamin D and VDR in the regulation of adiposity in vivo, animal models have been used. Reduced vitamin D signaling through global knockout of 1α-hydroxylase (CYP27B1) and VDR in mice leads to a lean phenotype when fed a high-fat diet or high-calcium rescue diet [3132]. In contrast, adipocyte-VDR null mice are fatter with sex-specific effects such that female but not male adipocyte-VDR knockout mice exhibit higher growth rates and increased visceral fat mass [33]. These studies imply that the lower fat mass in the global italicize VDR here and 1α-hydroxylase knockout models may not be due to the direct effects of vitamin D on adipocytes. However, transgenic mice overexpressing human VDR in adipose tissues are fatter and exhibit reduced energy metabolism without alterations in food intake [34], consistent with the phenotypes of the global knockout mice.
The lean phenotype and defective cellular adipogenesis in the VDR-null mice become apparent with age [32] and reduction in adipocyte size is observed in older (1 year) but not in young (21 days) mice [35]. Further, maternal vitamin D deficiency does not impact adipose tissue development in offspring [3536]. These results indicate that deficiency of VDR does not affect fat deposition during early development. Age-associated lean phenotype in the VDR-null mice may be due to the fact that 1,25(OH)2D3-VDR decreases energy expenditure through uncoupling process [13] and age-associated alopecia [35].
Role of vitamin D in adipose tissue function
Adipocytes store excess energy in the form of TAG and release them as FAs and glycerol when body energy demands increase during fasting or exercise. Adipose tissues are also endocrine organs secreting number of peptide hormones and cytokines including leptin, adiponectin and interleukin (IL)-6. By acting as both metabolic and endocrine organs, adipose tissues play crucial roles in energy homeostasis and alterations in adipose derived metabolic and endocrine products are thought to cause or exacerbate insulin resistance and other cardiometabolic diseases in obesity [20].
Effects of vitamin D on adipose metabolic functions
The amount of TAG storage in adipocytes is governed by the balance of lipid synthesis and breakdown. The role of vitamin D in adipocyte lipid metabolism is relatively unknown. An earlier study by Shi et al. [30] reported that 1,25(OH)2D3 stimulates calcium influx into adipocytes affecting lipid metabolism through the membrane bound VDR. Similarly, more recent studies showed that 1,25(OH)2D3 reduced TAG accumulation by increasing basal and adrenergically stimulated lipolysis [37] and decreasing de novo lipogenesis in 3T3-L1 adipocytes [38]. 1,25(OH)2D3 stimulates mRNA expression of several genes related to FA oxidation including CPT1A, PGC1α and PPARα as well as UCP1 [37] and rates of FA oxidation in 3T3-L1 adipocytes [38]. Overall, these data suggest that 1,25(OH)2D3 has catabolic effects in adipocytes decreasing lipid accumulation, which could potentially reduce the size of adipocytes (Fig. 4A). The non-genomic actions of vitamin D through calcium influx [30] may also explain the fact that calcium supplementation alone corrects most of the changes mediated by vitamin D deficiency in rats [39].
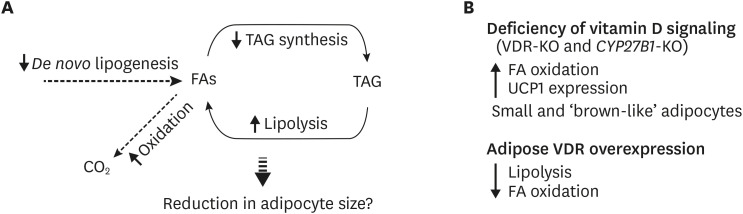 | Fig. 4Vitamin D regulation of adipose tissue metabolism. (A) Results from in vitro studies show that vitamin D increases lipolysis and FA oxidation while decreasing de novo lipogenesis. Combined, these may lead to the reduction in lipid accumulation and hence, reduction in adipocyte size. (B) Phenotypes of transgenic mouse models with alterations in vitamin D signaling are contradictory to the results from in vitro studies. Deficiency of vitamin D signaling lead to a leaner phenotype in mice with higher FA oxidation rates and UCP1 expression while VDR overexpressing mice exhibit the opposite phenotype.FA, fatty acid; UCP1, uncoupling protein 1; TAG, triacylglycerol; VDR, vitamin D receptor.
|
The phenotypes of VDR-null and overexpression transgenic mice do not support these catabolic effects of vitamin D on adipocyte lipid metabolism (Fig. 4B). Both VDR and 1α-hydroxylase (CYP27B1) knockout mice are lean and express higher levels of UCP1 in both white and brown fat and exhibit elevated levels of beta oxidation in WAT [3132]. Further, overexpression of human VDR in mouse adipose tissues reduced expression of genes involved in the regulation of FA transport, thermogenesis, and lipolysis and suppressed FA oxidation and lipolysis [34]. Some of these phenotypes may have been mediated by indirect catabolic effects of VDR on whole body energy homeostasis [1335].
Vitamin D is known to affect insulin actions and glucose metabolism in adipocytes. 1,25(OH)2D3 enhances insulin-stimulated AKT phosphorylation, GLUT4 translocation and glucose transport in 3T3-L1 adipocytes [4041]. In addition, vitamin D supplementation stimulates glucose uptake in adipose tissue of high fat diet-fed mice [42]. Results from transcriptome analysis in human adipocytes show that vitamin D increases oxidative stress [43], suggesting that it may impair insulin signaling pathway. Further studies of dissecting the role of vitamin D and VDR in human adipocyte metabolism are warranted.
Effects of vitamin D on adipose inflammation and endocrine function
Anti-inflammatory actions of vitamin D are well-known and most of studies showed that vitamin D decreases inflammation in adipose tissues. In both preadipocytes and adipocytes, 1,25(OH)2D3 suppresses expression levels of multiple cytokines including IL-6, IL-1β, IL-8, macrophage chemoattractant protein-1 and leptin [4041444546474849]. Further, 1,25(OH)2D3 stimulate adiponectin, an anti-inflammatory and insulin sensitizing adipokine, in 3T3-L1 adipocytes [41]. Contradictory results showing vitamin D increasing IL-6 and IL-8 in adipocytes [10] and leptin in mouse adipose tissues [50] have been also reported.
1,25(OH)2D3 decreases cytokines through the VDR in 3T3-L1 adipocytes [40] and in preliminary experiments, we found that 1,25(OH)2D3 suppresses proinflammatory cytokines through the VDR-dependent mechanisms in human adipocytes (unpublished observation, Nimitphong H and Lee MJ). Further, 1,25(OH)2D3 increases mRNA expression levels of dual specificity protein 10 and IκBα and inhibits NF-κB and p38 MAPK signaling pathways in 3T3-L1 adipocytes [40]. The proinflammatory actions of 1,25(OH)2D3 in 3T3-L1 adipocytes are known to be mediated through non-genomic actions of VDR in calcium signaling pathway [10].
Adipose tissues contain other cells including several types of immune cells and endothelial cells. In immune cells, vitamin D inhibits cytokine expression through the VDR-mediated suppression of proinflammatory NF-κB and MAPK signaling pathways [5152]. By suppressing chemokine production from multiple cell types, vitamin D-VDR blocks monocyte migration into adipose tissues, as demonstrated in mouse models in vivo [48]. In addition, 1,25(OH)2D3 also regulates the function of macrophages and other immune cell populations within adipose tissues [5354]. Overall, these results from in vitro studies and in vivo mouse models support that the anti-inflammatory actions of vitamin D in adipose tissues. By suppressing adipose tissue inflammation, vitamin D could improve systemic metabolism in obesity.
The in vivo effects of supplementation of vitamin D on serum levels of inflammatory cytokines in humans are controversial. Meta-analysis of vitamin D supplementation in obese and overweight subjects with mean baseline 25(OH)D levels from 12 to 32.6 ng/mL reported no significant changes in C-reactive protein (CRP), tumor necrosis factor-alpha (TNF-α) and IL-6 concentrations [55]. Another meta-analysis in 484 subjects with obese/overweight with or without type 2 diabetes also reported no significant effect of vitamin D supplementation on adiponectin and leptin levels [56]. On the contrary, a recent meta-analysis of 6 clinical trials with a mean baseline 25(OH)D of 24 ng/mL showed that vitamin D supplementation significantly increased serum leptin with a pool mean difference of about 5 ng/mL [57]. In a systematic review and meta-analysis, Yu et al. [58] reported that vitamin D supplementation significantly decreased CRP levels by 0.45 μg/mL without affecting TNF-α and IL-6, when studies in type 2 diabetes were exclusively considered. Therefore, different doses, duration and populations employed between studies may have contributed to the contradictory results.
Go to :
LOW VITAMIN D STATUS IN OBESITY AND METABOLIC DISEASES
Vitamin D status is assessed by serum 25(OH)D as levels of 1,25(OH)2D do not reflect vitamin D status [59]. Vitamin D status is influenced by diets, sun exposure, races, and genetic factors and the optimal serum 25(OH)D level for skeletal health is accepted to be above 20 ng/mL by the Institute of Medicine [60] or 30 ng/mL by the National Osteoporosis Foundation [61] and the International Osteoporosis Foundation [62].
Numerous studies have shown that low vitamin D status is associated with obesity and metabolic diseases [161718636465]. Several factors including lower vitamin D intake and sun exposure have been proposed to explain the lower vitamin D status in obesity. Others suggest that reduced serum levels are caused by both sequestration and volumetric dilution of vitamin D in larger amount of adipose tissues in obesity [6667]. Lower expression levels of CYP2J2 and CYP27B1 in adipose tissues [16] may also contribute to the vitamin D inadequacy in obesity.
Clinical effects of vitamin D supplementation on obesity
Interventional studies of vitamin D supplementation and adiposity measures and metabolic diseases remain inconclusive. We summarize results from human studies that had primary or secondary objectives to investigate the effect of vitamin D supplementation on obesity and metabolic diseases. In a systemic review and meta-analysis of 12 randomized controlled trials (RCTs) that had supplemented vitamin D without caloric restriction [68], Pathak et al. [68] showed that supplementation of various doses of vitamin D for 6 to 52 weeks resulted in neutral effects on standardized mean difference for body weight, fat mass, % fat mass or lean body mass. Another systemic review and meta-analysis investigated the effects of vitamin D supplementation alone (vitamin D3 in all except for one that used vitamin D2 and another that used alpha-calcidiol) or with calcium on adiposity measures [69]. This analysis included 26 RCTs and 42,430 participants with median treatment duration of 12 months and found no significant effects of vitamin D supplementation, compared to placebo or calcium control, on body mass index (BMI), body weight or fat mass. However, vitamin D plus calcium compared to placebo showed a small but significant reduction in body weight without effects on BMI or fat mass. The significant reduction in body weight was largely driven by the inclusion of the Women's Health Initiative Calcium/Vitamin D Supplemental Trial [70]. Of note, only changes in body weight, but not BMI and fat mass, were reported in the trial. An analysis for a dose-response effect by vitamin D3, from < 1,000 IU/day to > 4,000 IU/day, revealed no effect on any of the adiposity outcomes in any dosage groups [69].
Since those 2 reviews were published, there have been 6 RCTs that examined the effects of vitamin D supplementation on adiposity [717273747576]. Results from these studies also do not support the role of vitamin D in reducing adiposity. Further, in a bi-directional genetic approach using Mendelian randomization to limit confounding, Vimaleswaran et al. [77] showed that although a higher BMI was causally related to lower 25(OH)D, vitamin D deficiency was not a causal factor for the development of obesity. Therefore, RCTs of vitamin D supplementation have not provided evidence of cause-effect relation between vitamin D status and body weight control.
Clinical effects of vitamin D on metabolic disorders
A meta-analysis consisted of 1,181 individuals with BMI > 23 kg/m2 and normal or impaired fasting glucose levels showed that vitamin D and/or calcium supplementation has no significant effect on fasting glucose and insulin levels as well as homeostatic model assessment-insulin resistance (HOMA-IR) index [78]. Importantly, the results were similar when the supplemented doses (low dose: 125–2,000 IU/day vs. high dose: 3,571–4,000 IU/day), durations of supplementation (short: until 15-weeks vs. long: > 15 weeks) or baseline of 25(OH)D concentrations (deficiency: < 50 nmol/L [20 ng/mL] vs. insufficiency: 52.5–72.5 nmol/L [21–30 ng/mL]) were taken under consideration in subgroup-analysis. Similarly, in a recent systematic quantitative review on findings from meta-analyses, Rejnmark et al. [79] also showed null-findings in the risk of cardiometabolic diseases with vitamin D supplementation. A meta-analysis of 24 RCTs, however, showed that a significant reduction in HbA1c, fasting plasma glucose and HOMA-IR following vitamin D supplementation in type 2 diabetic patients [80]. The authors suggest that a minimum dose of 4,000 IU/day of vitamin D which brings serum 25(OH)D values to higher than 100 nmol/L (40 ng/mL) may be recommended as an adjunct therapy to improve glycemic measures in type 2 diabetic patients. In addition, combined supplementation of calcium plus vitamin D for eight weeks significantly improved glucose and lipid metabolism in overweight to obese, vitamin D-deficient women with polycystic ovary syndrome [81]. Differences in study designs including doses, durations, baseline vitamin D status and study subjects may explain contradictory results and more studies investigating in a specific group of participants with or without true vitamin D deficiency are needed.
Go to :
Conclusion
Adipose tissues act as a storage or buffering site of vitamin D but also participate vitamin D metabolism. Vitamin D affects new fat cell formation as well as metabolic and endocrine functions of adipose tissues. Results from cell culture and mouse models assessing the role of vitamin D in adipogenesis are inconsistent. Both suppression and promotion of adipogenesis have been reported depending on the cell culture models and conditions used. Both CYP27B1 and VDR knockout mice are lean while transgenic mice overexpressing human VDR in adipocytes are obese, supporting the positive actions of vitamin D-VDR on adipogenesis in vivo. However, results from more careful studies suggest that the phenotypes of the transgenic mice are due to vitamin D actions on systemic energy metabolism, rather than direct effects on adipocyte development. Although vitamin D actions on adipose metabolism are relatively unknown, several studies show that it regulates lipolysis and lipid synthesis and improve insulin signaling pathways. In addition, vitamin D may improve adipose tissue inflammation. These results suggest that vitamin D may ameliorate adipose tissue dysfunctions, linking low vitamin D status to metabolic disease in obesity as shown in many association studies. Whether maintenance or restoration of vitamin D status has beneficial effects on systemic metabolism is not known and more studies are warranted to establish a causal relationship. Further elucidation of the role of vitamin D and mechanisms through which vitamin D regulates adipose biology may lead to discovery of therapeutics to improve metabolic health in obesity.
Go to :
 XML Download
XML Download