

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Hepatic steatosis is the most important cause of chronic disease. It is characterized by excessive fat accumulation in the liver, which can eventually lead to hepatocellular carcinoma [1]. The prevalence of hepatic steatosis among adults with obesity, diabetes, and hyperlipidemia in Western countries is 20–30%, similar to that in South Korea [23]. Hepatic steatosis is more likely to occur in women than in men, particularly among those aged 56–60 years, corresponding to the post-menopausal transitional period [4].
During menopause, there is an age-related decline in ovarian function and estrogen levels. Recently, the relationship between menopause and histopathological alterations in the liver, which is the primary target organ of estrogen, has attracted attention [5]. There is increasing evidence that estrogen regulates lipid metabolism, and its deficiency following menopause is associated with the occurrence of hepatic steatosis [6]. In fact, estrogen replacement therapy reduces the risk of fatty liver [7]. In menopause, a loss of the liver's capacity to oxidize fatty acids occurs, along with an increase in lipogenesis, resulting in excessive fat accumulation in the liver [8]. Therefore, menopausal women should be considered targets for clinical interventions for further investigations of hepatic steatosis.
Rice (Oryza sativa L.) is an important cereal crop consumed by more than half of the world's population as a staple food [9]. Rice bran, the byproduct of rice milling, contains an array of phytochemicals, such as phenolics, flavonoids, vitamins, amino acids, and phytosterols. Various studies have indicated that rice bran has diverse health effects, including antioxidant, anti-inflammatory, cholesterol-lowering, anti-hyperlipidemic, and anti-diabetic activities [10111213]. In addition, several clinical studies reported that the consumption of stabilized rice bran, which is rice bran inactivated by the lipase enzyme through heating, decreases fasting glucose, total cholesterol (TC), low-density lipoprotein cholesterol, apolipoprotein B, and triglyceride (TG) levels, and increases insulin in the sera of patients with diabetes [1415]. In particular, γ-oryzanol, which is a major compound in rice bran, reduced the bone mineral density loss associated with ovariectomized (OVX)-induced estrogen deficiency in rats via γ-amino butyric acid B receptors [16]. Moreover, it ameliorates lipopolysaccharide-induced cognitive and memory impairments in mice by promoting hippocampal antioxidant and anti-inflammatory molecular responses [17]. Despite the diverse pharmacological activities of rice bran, its effects and underlying molecular mechanisms in protecting against hepatic steatosis induced by estrogen deficiency are unclear. In the present study, we hypothesized that a standardized rice bran extract (RBS) impacts hepatic steatosis induced by estrogen deficiency by altering lipogenesis-related genes. Therefore, the protective effects of RBS on the liver were investigated in the livers of OVX rats, animal models of human menopause, as well as in oleic acid-treated HepG2 cells [18].
MATERIALS AND METHODS
Preparation of RBS
The RBS (Lot No. SD-RB-002) standardized with 0.45% γ-oryzanol was obtained from S&D Co., Ltd. (Cheongju, Korea) as previously described [192021]. Briefly, rice bran (O. sativa L.) was extracted in an ethanol/water solution at 40°C for 8 h. The ethanol extract was filtered, concentrated, and spray-dried for storage. The yield of RBS was 29.3%. HPLC analysis was performed using a LiChrospher 100 Diol column (250 × 4.0 mm, 5 µm; Merck Millipore, Billerica, MA, USA). The solvent systems used for separation were an isocratic elution (A: 0.1% acetic acid in cyclohexane, and B: 0.1% acetic acid in ethyl acetate) in a ratio of 85:15 (v/v). The injection volume was 10 µL and the column eluent was monitored at UV 315 nm; chromatography was performed at 30°C with a flow rate of 0.5 mL/min. The HPLC quantitative analysis was replicated three times. A representative chromatogram of the γ-oryzanol reference and its corresponding peak in the RBS is presented in Fig. 1.
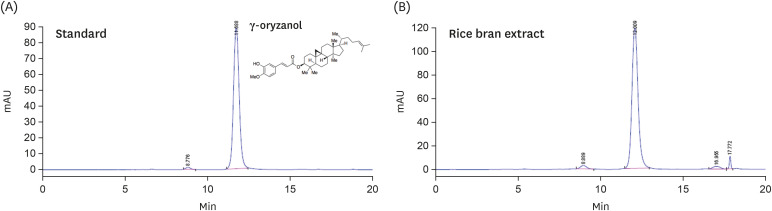
Cell culture and oil red O (ORO) staining
HepG2 cells were obtained from ATCC (Manassas, VA, USA). The cells were maintained in DMEM supplemented with antibiotics (100 U/mL penicillin A and 100 U/mL streptomycin) and 10% fetal bovine serum. The cells were treated with or without samples in oleic acid (200 µM) for 24 h. Lipid droplets were visualized and subsequently quantified by ORO staining. The cells were fixed in 10% formaldehyde for 1 h, washed with 60% isopropanol, and mixed with ORO solution. Red-stained lipid droplets were observed with a light microscope. To quantify lipid accumulation, ORO was eluted with 100% isopropanol, and then, the optical densities of the eluates were measured using a spectrophotometer at 500 nm.
Animal studies
Female sham-operated and OVX rats (Sprague-Dawley, 6 weeks of age) were purchased from SLC (Shizouka, Japan). The rats were maintained at a temperature and humidity of 21–25°C and 50–60%, respectively, and under a 12-h:12-h light:dark cycle with free access to water and food. Thirty rats were subjected to the removal of the bilateral ovaries (OVX), and 10 rats were subjected to incision and suturing without ovary removal (SHAM group). After a 2-week recovery period, the OVX rats were randomly divided into 3 groups (n = 10 per group) as follows: OVX rats fed a normal diet (ND; OVX group), OVX rats fed an ND containing 17β-estradiol (E2, 10 µg/kg body weight; OVX+E2 group), and OVX rats fed an ND containing RBS (500 mg/kg body weight; OVX+RBS group) for 16 weeks. The dosage of RBS and E2 were chosen based on our preliminary study and previous reports [202223]. This dosage was converted from dose per kg body weight to % of diet considering daily intake and body weight of rats. Finally, rats were fed an ND containing 0.00002% E2 or ND containing 1% RBS. The ND was prepared by supplementing the basal American Institute of Nutrition-93G diet (Research Diets, New Brunswick, NJ, USA). The experimental diets were prepared by mixing RBS or E2 into the AIN-93G diet, shown in Supplementary Table 1.
Body weight was measured once per week, and food intake was measured 3 times per week. After feeding the rats with the experimental diets for 16 weeks, we collected blood from the retro-orbital sinus in the fasted state. The sera were separated and stored at −80°C until analysis. The liver and adipose tissues were removed, weighed, and stored at −80°C. All animal experiments were approved by the Korea Food Research Institutional Animal Care and Use Committee (permission number: KFRI-M-16056).
Serum biochemical analysis
Serum TC, TG, high-density lipoprotein-cholesterol (HDL-C), free fatty acid (FFA), and glucose levels were measured using commercial enzyme kits (Shinyang Chemical Co., Ltd., Seoul, Korea). Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were measured using commercial reagent kits (Young-Dong Diagnostics, Yongin, Korea).
Hepatic lipid analysis
Hepatic total lipids were extracted as described by Folch et al. [24]. Hepatic TG and TC were measured using a commercial enzyme kit (Shinyang Chemical Co., Ltd.).
Histological analysis
Liver tissue was removed from the same part of the left lobe of each rat and fixed with 4% formaldehyde in phosphate-buffered solution. The paraffin-embedded liver samples were sliced into 5-µm-thick sections and stained with hematoxylin and eosin (H&E). Stained sections were analyzed by light microscopy (BX50, Olympus, Tokyo, Japan), and digital images were captured.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
The expression of the sterol regulatory element-binding protein 1 (SREBP1), acetyl-CoA carboxylase 1 (ACC1), fatty-acid synthase (FAS), stearoyl-CoA desaturase 1 (SCD1), and glycerol-3-phosphate acyltransferase 1 (GPAT) genes was analyzed by qRT-PCR. The primers used in this study are shown in Table 1. Total RNA was extracted from the liver tissue using a total RNA isolation kit (Macherey-Nagel, Düren, Germany) according to the manufacturer's protocol. The cDNA was synthesized using ReverTra Ace qPCR RT Master Mix (Toyobo, Osaka, Japan). qRT-PCR was performed with SYBR Green Master Mix (Toyobo) and forward/reverse primers using a QuantStudio 6 Flex RT PCR System (Applied Biosystems, Foster City, CA, USA). The relative gene expression levels were normalized to that of β-actin.

Table 1
Primer sequences
ACC1, acetyl-CoA carboxylase 1; FAS, fatty acid synthase; GPAT, glycerol-3-phosphate acyltransferase; SCD1, stearoyl-CoA desaturase 1; SREBP1, sterol regulatory element-binding protein 1.
![]()
RESULTS
Effect of RBS on lipid accumulation in oleic acid-induced HepG2 cells
To investigate the effect of RBS on oleic acid-induced intracellular lipid accumulation, HepG2 cells were treated with 200 µM oleic acid and RBS (100 and 250 µg/mL) or γ-oryzanol (1 and 10 µM). As shown in Fig. 2, oleic acid-treated HepG2 cells showed a significant increase in lipid droplets, whereas untreated control cells did not. However, the development of lipid droplets induced by oleic acid was significantly decreased by RBS or γ-oryzanol treatment.
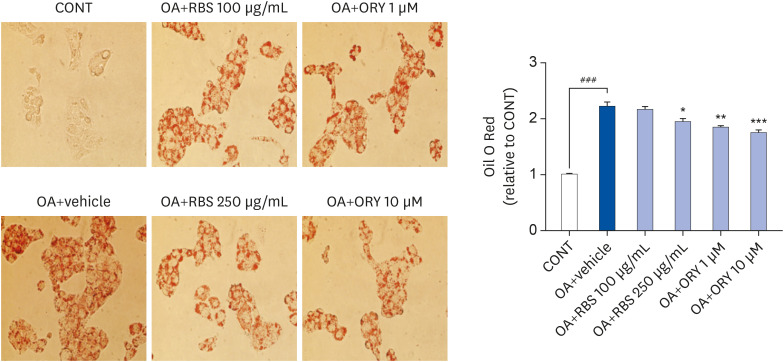
Fig. 2
Effects of RBS and γ-oryzanol on intracellular lipid accumulation in OA-stimulated HepG2 cells. After treatment with 200 µM OA with or without RBS and γ-oryzanol for 24 h, the cells were stained with ORO solution and intracellular lipid accumulation was visually observed under a light microscope (magnification, ×200). Data are expressed as mean ± SD.
RBS, rice bran extract; ORO, oil red O; CONT, not-treated cells; OA, oleic acid; ORY, γ-oryzanol; RBS, rice bran extract.
###P < 0.001 compared to untreated cells. *P < 0.05, **P < 0.01, and ***P < 0.001 compared to OA only treated cells.
![]()
Effect of RBS on body and organ weights
Compared to those in the SHAM group, the OVX group showed significant increases in the final body weight, body weight gain, and fat pad weight. However, the final body weight, body weight gain, and fat pad weight in the RBS group were lower than those in the OVX group, although the differences were not significant. We also found that liver weight was significantly higher in OVX rats than in SHAM rats, and this increase was significantly attenuated by RBS supplementation (Table 2).

Table 2
Effects of RBS on body weight and organ weights in OVX rats fed an experimental diet for 16 weeks
Data are expressed as mean ±SD.
SHAM, sham-operated controls; E2, 17β-estradiol; OVX, ovariectomized; RBS, rice bran extract.
#P < 0.05, ###P < 0.001 compared to the SHAM group. **P < 0.01, ***P < 0.001 compared to the OVX group.
![]()
Effect of RBS on blood parameters
As shown in Table 3, OVX rats exhibited significantly higher serum TG and FFA levels than the SHAM group. In contrast, RBS supplementation restored serum TGs and FFAs to levels similar to those of the SHAM group. However, TC, HDL-C, ALT, and AST levels did not differ significantly among the experimental groups.
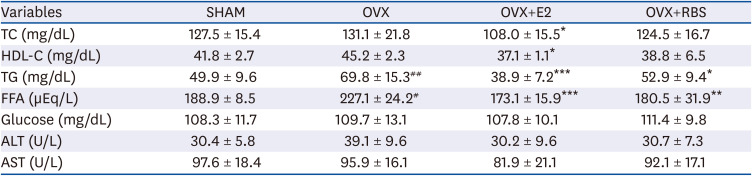
Table 3
Effect of RBS on fasting serum lipids levels in OVX rats fed an experimental diet for 16 weeks
Data are expressed as mean ±SD.
RBS, rice bran extract; OVX, ovariectomized; SHAM, sham-operated controls; E2, 17β-estradiol; TC, total cholesterol; HDL-C, high-density lipoprotein cholesterol; TG, triglyceride; FFA, free fatty acids; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
#P < 0.05, ##P < 0.01 compared with SHAM group. *P < 0.05, **P < 0.01, and ***P < 0.001 compared with OVX group.
![]()
RBS attenuates hepatic lipid accumulation
Next, we investigated whether RBS protects against OVX-induced hepatic steatosis. As shown Fig. 3, histological analysis showed that OVX rats developed hepatocellular micro- and macrovesicular vacuolation, but these alterations were ameliorated by RBS (Fig. 3). Furthermore, increases in hepatic total lipids, TG, and TC levels induced by OVX were significantly attenuated by RBS (45.5%, 49.3%, and 22.3% reductions compared to levels in the OVX group, respectively), similar to those in the OVX+E2 group (Table 3). These data indicate that RBS exerts inhibitory effects on hepatic steatosis.
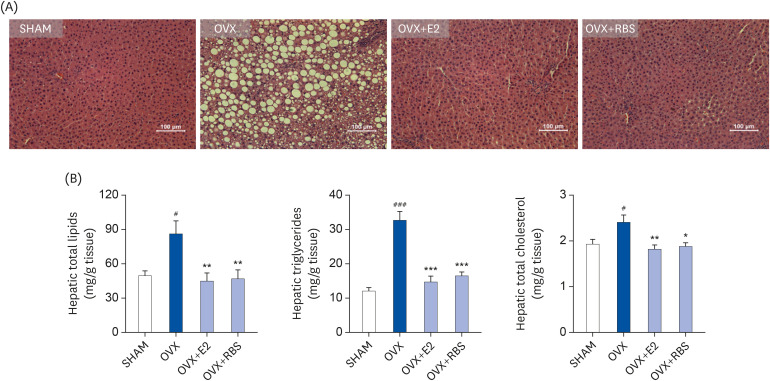
Fig. 3
Effect of RBS on hepatic lipid levels in OVX rats. (A) Histological changes (original magnification, ×200) and (B) hepatic lipid, triglyceride, and total cholesterol levels in rats. Data are expressed as mean ± SD.
RBS, rice bran extract; OVX, ovariectomized; E2, 17β-estradiol; SHAM, sham-operated controls.
#P < 0.05 and ###P < 0.001 compared to the SHAM group. *P < 0.05, **P < 0.01, and ***P < 0.001 compared to the OVX group.
![]()
RBS ameliorates expression of lipogenic genes
It is possible that RBS reduces fat accumulation in hepatocytes by regulating the expression of genes involved in lipogenesis. To test this hypothesis, we analyzed the hepatic expression of genes associated with lipogenesis by qRT-PCR. As shown in Fig. 3, the mRNA expression levels of lipogenic genes, including SREBP1, ACC1, FAS, SCD1, and GPAT, were increased in the livers of OVX rats compared to those in the SHAM group. In contrast, RBS supplementation decreased the expression levels of these genes (Fig. 4). These data suggest that RBS suppresses OVX-induced fatty liver by downregulating the expression of lipogenesis genes.
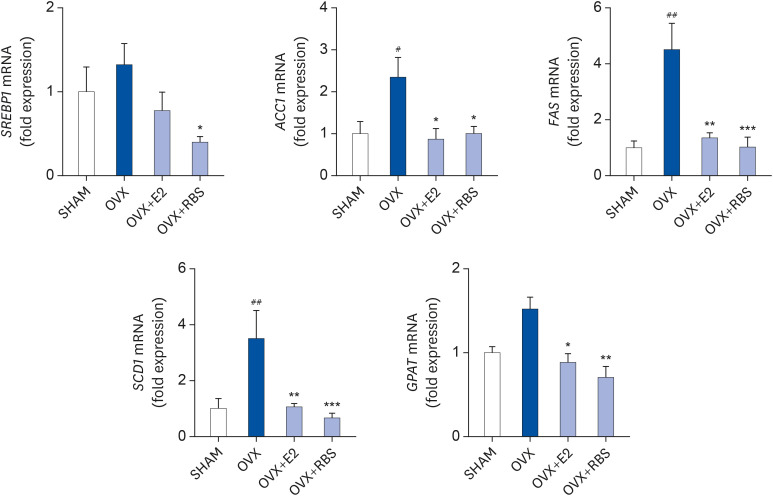
Fig. 4
RBS suppresses the mRNA expression of lipogenic genes. Relative mRNA levels were measured by qRT-PCR and normalized against that of β-actin. Data from three independent experiments are presented as the mean ± SD.
RBS, rice bran extract; qRT-PCR, quantitative reverse transcription polymerase chain reaction; SHAM, sham-operated controls; OVX, ovariectomized; E2, 17β-estradiol; SREBP1, sterol regulatory element-binding protein 1; ACC1, acetyl-CoA carboxylase 1; FAS, fatty acid synthase; SCD1, stearoyl-CoA desaturase 1; GPAT, glycerol-3-phosphate acyltransferase.
#P < 0.05, ##P < 0.01 compared to the SHAM group. *P < 0.05, **P < 0.01, and ***P < 0.001 compared to the OVX group.
![]()
DISCUSSION
Postmenopausal women exhibit more frequent development of hepatic steatosis than premenopausal women. For example, aromatase-knockout mice, a useful model for examining the role of estrogen, exhibit hyperlipidemia and hepatic steatosis [26]. In addition, OVX-induced estrogen decreases significantly increase serum lipoprotein levels and hepatic fat accumulation, whereas E2 treatment completely restores normal levels [27]. Heine et al. [28] reported that estrogen receptor-knockout mice exhibit greater fat masses and lower energy expenditures than wild-type mice. Estrogen has been reported to play a role in liver fat accumulation; thus, estrogen deficiency leads to a disturbance of lipid regulatory mechanisms in the liver [29]. As estrogen levels decrease, this lipid accumulation might be the result of several factors, including alterations in the uptake or export of fatty acids, changes in the mitochondrial oxidation of fatty acids, and reduced export of TGs [30]. Thus, fat accumulation in the liver is of critical concern in postmenopausal women. Therefore, we investigated the protective effects of RBS on HepG2 cells and OVX-induced hepatic steatosis in rats. In the present study, we found that lipid accumulation was significantly increased in oleic acid-treated HepG2 cells. RBS and γ-oryzanol, a major component of RBS, exerted an inhibitory effect on oleic acid-induced lipid accumulation in HepG2 cells. In agreement with the in vitro data obtained using HepG2 cells, we found that OVX caused hepatic fat accumulation, whereas RBS supplementation suppressed OVX-induced increases in hepatic lipid accumulation and lipid levels. Moreover, RBS reduced serum TG and FFA levels. RBS supplementation could have induced this reduction in circulating lipid levels by decreasing lipid transport to the liver, thereby preventing hepatic lipid accumulation. Surprisingly, in our study, the serum AST and ALT levels, used as indicators of hepatic damage [31], did not differ among the experimental groups. Charatcharoenwitthaya et al. [32] reported that the level of hepatic enzymes is an insensitive tool to follow histological changes in the liver among patients with nonalcoholic fatty liver disease. Moreover, in animal studies comparing OVX and SHAM mice, serum markers of liver function, ALT or AST, showed no significant differences between both groups, although fat accumulation was observed in the OVX animals [3334]. Thus, our results suggest that the effects of RBS on circulating lipid profiles in serum can be attributed to improved hepatic lipid accumulation.
In the present study, RBS altered hepatic lipid metabolism and the expression of genes involved in these processes. Liver fatty acid synthesis might result in hepatic steatosis and is mostly regulated by well-known transcription factors, such as SREBP1. SREBP1 is the key regulator of hepatic fatty acid and TG synthesis [35], and reducing its expression is thought to improve hepatic steatosis. SREBP-1c activates ACC1, FAS, SCD1, and GPAT, which are responsible for lipogenesis in the liver. ACC1, the enzyme that carboxylates acetyl-CoA to malonyl-CoA, is utilized by FAS in the synthesis of fatty acids [36]. A previous study showed that SCD1, a central lipogenic enzyme catalyzing the synthesis of monounsaturated fatty acids, -deficient mice display reduced lipid synthesis and enhanced lipid oxidation and insulin sensitivity in the liver [37]. Moreover, the human fatty liver is characterized by increased hepatic SCD1 levels and lipogenic activity [38]. GPAT catalyzes the first step in the synthesis of TGs and is recognized as contributing to lipogenesis in the liver [39]. Interestingly, GPAT has been suggested as a potential therapeutic target for dyslipidemia and obesity [40]. As expected, our results showed that the expression levels of the hepatic lipogenic genes SREBP1, ACC1, and FAS were increased in the livers of OVX rats, which is supported by previous studies showing that lipogenesis-related protein expression in the liver is dramatically increased in OVX animal models [4142]. Based on our results, SREBP1-induced expression of FAS and ACC1 mRNA might have affected the production of fatty acids. These fatty acids are used as a source for TG synthesis by SCD1 and GPAT, thereby triggering lipid accumulation in hepatocytes. However, in the present study, RBS supplementation notably inhibited the mRNA expression of these lipogenic genes. Our findings suggest that RBS ameliorates OVX-induced hepatic lipid accumulation by regulating genes involved in lipogenesis. Hao et al. [43] reported that a pre-germinated brown rice extract decreases the protein expression of SREBP1, SCD1, and FAS in the livers of high-fat diet (HFD)-fed mice. Moreover, the typical hepatic steatosis observed in the livers of ApoE−/− mice fed an HFD was reduced by rice bran enzymatic extract [44]. These effects could have been the result of various bioactive compounds (phytic acid, γ-oryzanol, GABA, phenolics, and anthocyanins) present in the rice bran [45]. Among them, γ-oryzanol, which consists of a mixture of ferulic acid esters of sterols, has been suggested to reduce circulating levels of liver lipoproteins in obese rats [46]. Furthermore, γ-oryzanol alleviates high-fat and high-fructose diet-induced hepatic steatosis through the inhibition of intracellular TG and TC accumulation in male SD rats and downregulates the expression of the lipogenic genes FAS, SCD1, ACCα, and ACCβ in HepG2 cells [47]. In the study by Wang et al. [48], γ-oryzanol was found to ameliorate HFD-induced obesity in mice and also reduced obesity-induced chronic inflammation in the hepatic tissues of mice. The authors indicated that γ-oryzanol-mediated inhibition of nuclear factor-κB activation might have participated in both anti-inflammation and lipid-lowering effects. Song et al. [49] reported that phytosterol esters attenuate hepatic steatosis in rats fed a HFD. Although not all components of RBS have been identified, we measured the content of γ-oryzanol in the RBS as 4.5 mg/g extract. Additionally, γ-oryzanol significantly inhibited lipid accumulation in oleic acid-treated HepG2 cells. Thus, various components of RBS likely contributed to its hepatoprotective effects. Collectively, our results indicate that RBS inhibits increases in serum TG and FFA levels and hepatic lipid accumulation by downregulating SREBP1-mediated activation of lipogenic genes. This study provides a basis for the use of RBS as a supplement for the treatment or prevention of fatty liver in postmenopausal women.
 XML Download
XML Download