

 PDF
PDF Citation
Citation Print
Print



서 론
2003년 인간게놈프로젝트(human genome project) 완료 이후 많은 과학자들은 인체 내에 존재하는 미생물(microbiota)의 유전 정보 분석에 관심을 가지게 되었다.1 인체는 미생물이 공생하는 복잡한 생태계이다.2 미생물은 체내 대사 작용에 관여하고 유입된 외부 인자와의 상호작용을 포함한 다양하고 중요한 역할을 한다. 이 때문에 많은 연구자들은 체내에서 관찰되는 인간세포 이외의 미생물 유전체를 ‘세컨드 게놈(second genome)’ 혹은 ‘마이크로바이옴(microbiome)’이라 명명하며 그 유전 정보들을 분석하기 시작하였다.3 본격적으로 해당 연구가 촉발된 것은 2007년 국가별로 자국민에 대한 질병을 일으키는 미생물의 유전 정보를 분석하고 이를 공유하기 위하여 여러 나라에서 모여 ‘국제인간마이크로바이옴 컨소시엄(International Human Microbiome Consortium)’을 조직하면서부터이고 한국은 2011년 5월 8번째 회원국으로 가입하였다.4 최근 임상 영역에서는 장내 미생물의 유전 정보를 진단 및 치료 요법에 사용하려는 시도들이 지속되고 있고, 장내 미생물 환경의 조절요법에 대한 연구들이 진행 중이다. 췌담도계를 통하여 장관 내로 배출되는 담즙이나 췌장외분비액은 장내 화학 환경(chemical environment)에 직접적인 영향을 미치는 것들로 미생물들의 생태계에 영향을 준다. 또한 담도계 질환의 많은 원인을 차지하는 감염병의 원인균들이 사실은 장내 미생물로부터 유래한 것이며, 강력한 지질과 단백 분해 물질을 분비하는 췌장도 미생물이 존재하는 장기임이 알려졌다.5-8 이러한 미생물의 존재와 함께 악성 종양과 자가면역 질환을 포함한 여러 질환자에게서 장내 미생물의 변화가 관찰되어 질병과 무관하지 않을 것으로 추정된다.6,9-13 본고에서는 장내 미생물의 조절자로서 췌담도계의 기능에 대하여 알아보고, 장내 미생물과 관련된 췌담도계 질환에 대하여 알아보고자 한다.
Go to : 
본 론
1. 장내 미생물
그동안 장내 미생물들이 인체에 미치는 영향에 대하여 많은 연구들이 진행되어 왔고, 대사 작용 및 질환의 발생에 대한 것들이 규명되었다. 염증성 장질환(inflammatory bowel disease), 만성 피로 증후군(chronic fatigue syndrome), 비만, 제2형 당뇨, 암과 같은 질환이나 자폐증, 파킨슨병(Parkinson’s disease)과 같은 정신•신경계 질환까지도 포함한 많은 질병들 과 연관이 있음이 보고되었다.14-26 그중 자폐증에 대한 연구는 2016년 Buffington 등24에 의하여 진행된 동물모델 실험 결과로 많은 관심을 모았던 것이 2019년 Kang 등25에 의한 임상실험 결과까지 보고되면서 연구의 대상을 넓힌 좋은 예라고 할 수 있다.
장내 미생물 조성은 개인, 가족, 인종, 성별, 나이, 식습관, 지역, 생활방식 등 다양한 원인에 의하여 역동적으로 변화한다. 그러나 장내 미생물 환경을 이루는 주요 기능성 유전자들 (core microbiome)은 개체 특성과 환경 조건의 차이에도 상당 부분 일치한다.27,28 이는 인간을 포함한 같은 동물세포 DNA에 공통된 부위가 존재하듯 체내 존재하는 미생물도 개인간 차이가 있지만 같은 분류의 동물끼리 공통된 종류의 미생물이 체내 존재한다는 것이다. 체내 존재하는 미생물 유전정보가 종-특이성을 가지고 있다는 것은 그들의 생태계에 영향을 미치는 공통된 인자가 있다는 것이다. 해부학적 차이와 장내 내인성 항생 물질, 섭취하는 음식물의 종류가 가장 주된 원인으로 추정된다.
인간을 포함한 포유류에서 장내 미생물은 다양한 환경적 노출과 숙주의 영향을 받아 점차 그 구성이 복잡•다양화되는데, 성인이 되면서 점차 안정화된다.29 성인의 대장 내에는 비건조대변에 2-5×1011/g의 미생물이 존재하는 등 인간의 전체 장관 내에는 약 1,000문(門, phylum)의 미생물이 존재한다. 분류학 상피르미쿠테스(Firmicutes), 박테로이데테스(Bacteroidetes), 프로테오박테리아(Proteobacteria), 아시네토균(Acinetobacteria), 푸소박테리아(Fusobacteria)와 베루코마이크로비아(Verrucomicrobia) 계열 등의 6가지가 대부분을 차지하며, 그중 피르미쿠테스와 박테로이데테스가 90% 이상을 차지한다고 알려져 있다.17
Donaldson 등30이 작성한 장내 미생물 지도(gut biogeography)를 보면 대부분의 많은 미생물들이 대장에 존재하며, 소장에서도 원위부로 갈수록 개체수가 증가함을 알 수 있다(Fig. 1).30 이는 근위부 소장 내에 형성된 위산에 의한 산성 환경과 고농도의 산소, 췌장효소 및 담즙산 등에 영향을 받는 것으로 추정된다. 소장 내부는 내인성 항생 물질에 대한 저항력이 있고, 장관 내로 전달되는 단순당(simple carbohydrate) 흡수에 있어 다른 미생물이나 숙주에 대한 경쟁력이 있는 능률이 좋은 혐기균이 지배하고 있다. 담즙산은 미생물 억제 능력이 있는 강력한 내인성 항균 물질로 대부분 소장에 존재하고 말단회장 이전에 재흡수되어 대장에는 매우 소량만 존재하기 때문에 대장에는 다량의 미생물이 존재하며, 균주의 종류도 소장과 차이를 보인다. 또한, 장관의 단면상 분포를 보면 점막과의 거리가 가까워 산소 확산 농도가 높고 점막과의 면역학적 상호작용이 많이 일어나는 주름 사이와 산소 확산 농도가 낮고 면역학적 상호작용이 적은 내강 중심부의 균주 차이를 확인할 수 있다.
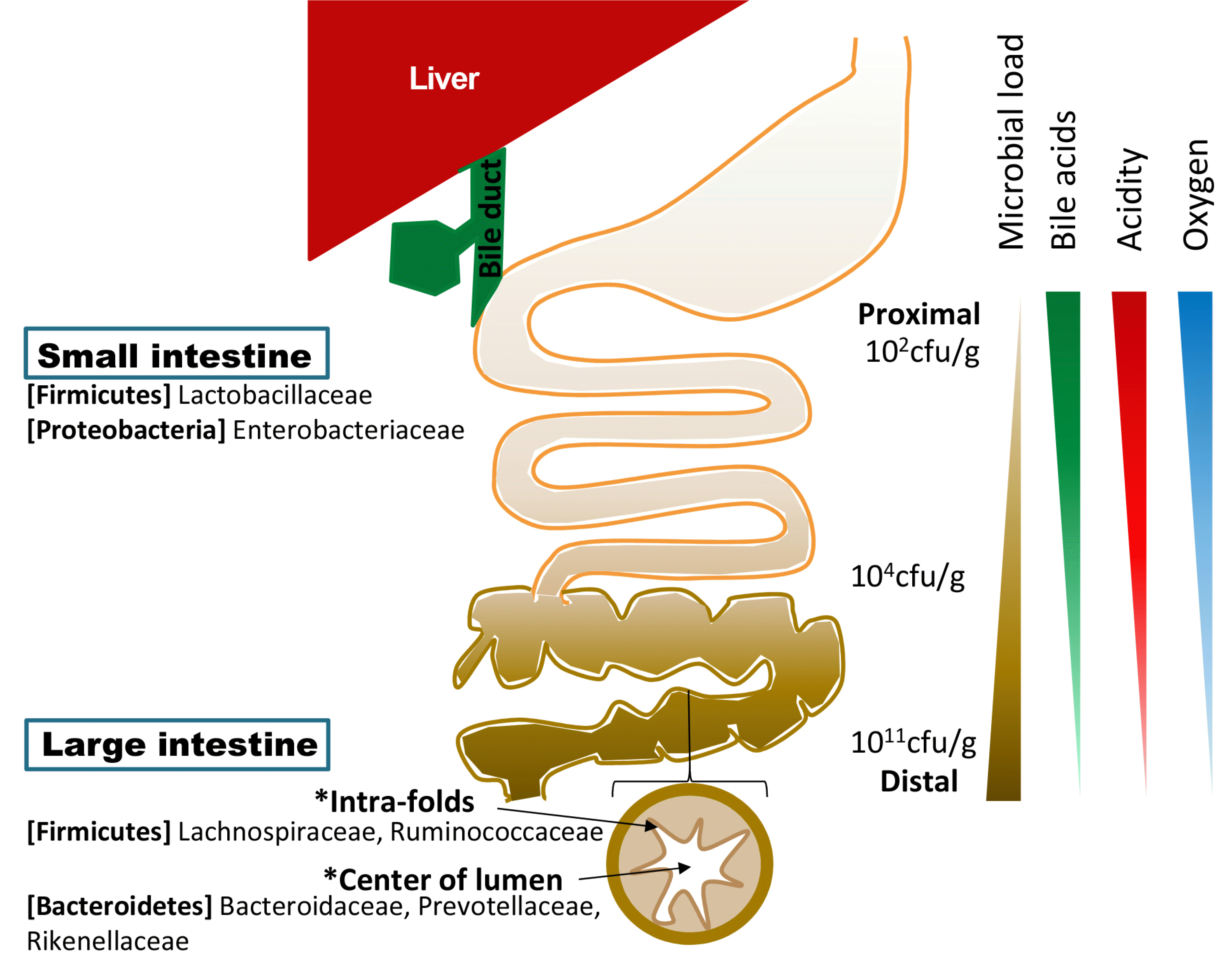
2. 장내 미생물 조절자
미생물에 대한 내인성 항생 물질 중 가장 중요한 조절자는 담즙산(bile acids)이다. 이는 콜레스테롤로부터 합성되어 담도계를 통하여 담즙에 섞여 십이지장으로 배출되고 대부분이 말단회장 이전에 재흡수된다. 인간의 간에서 고전적 경로와 대체 경로를 통하여 생성된 콜산(cholic acid)과 케노데옥시콜산(chenodeoxycholic acid)은 위장관으로 분비되어 일부가 장내 미생물에 의한 탈수 소화 과정을 통하여 2차 담즙산인 데옥시콜산(deoxycholic acid)과 리토콜산(lithocholic acid)으로 변화하게 된다. 재흡수된 담즙산은 장-간순환(enterohepatic circulation)을 통하여 간으로 이동 후 글라이신(glycine)과 타우린(taurine)으로 포합(conjugation)되고 담도계을 통하여 재분비된다. 이로 인하여 장관 내에는 매우 다양한 종류의 담즙산이 존재하게 된다. 담즙산은 장관 내로 분비되어 지방질을 흡수하는데 도움을 주기도 하고 체내 콜레스테롤을 배출하는 수단으로 이용되기도 한다. 담즙산은 장내 미생물에 대한 두 가지 억제 효과를 가진다.31-33 하나는 미생물 세포에 대한 직접적인 독성이고 가장 대표적인 것이 세포벽에 대한 계면활성제 성질(detergent property)이 미치는 직접적인 항균 효과이며,34-37 이외에 DNA 손상,38-40 단백 변성,34,41 철과 칼슘의 흡착 등이 영향을 미친다.38,42-44 다른 하나는 장과 간담도의 farnesoid X receptor (담즙산 핵 수용체[FXR], NR1H4)가 매개하는 간접적 억제 효과이다.45-49 FXR은 음성되먹임을 통하여 담즙산의 생산 및 장간순환량을 조절하며 지방과 포도당의 대사와 관련된 다양한 기능을 수행하는 핵 내 수용체 중 하나이다.50,51 담즙산의 FXR을 통한 신호 전달은 장내 면역 체계를 조절하고49 점막 장벽을 유지하며46 angiogenin gene Ang1과 the nitric oxide synthase gene iNos의 활성화 과정을 통하여45,46,52 미생물을 억제하고 장벽을 보호한다. 또한 담도와 장벽에서 다른 담즙산 수용체인 vitamin D receptor와 함께 cathelicidin과 같은 항균 펩타이드(antimicrobial peptides, AMPs)를 합성한다.53,54 소장 내에서 담즙산은 지속적으로 흡수되어 원위부로 갈수록 농도가 낮아지는데 근위부에서는 고농도 담즙산에 의한 직접적인 독성이 미생물을 억제하는 반면에 원위부에서는 FXR이 매개하는 간접 경로를 통한 억제의 비중이 높아진다.46,55,56
담즙산은 소수성 성질과 친수성 성질을 동시에 가지고 있는 양극성의 스테로이드 구조이고 결합된 잔기(residue)의 종류와 위치에 따라 다른 성질을 가진다. 미생물에 대한 직접적인 억제력은 소수성의 정도가 큰 담즙산일수록 그 정도가 크다고 알려져 있고, FXR에 대한 친화도가 클수록 미생물 억제능이 크다. 이러한 성질의 조합은 각 담즙산마다 모두 달라서 종류별 미생물들에 대한 억제 효과가 매우 다양하고 각 미생물들도 형태나 성질에 따라서 각 담즙산들에 대한 반응이 다양하다(Fig. 2).57
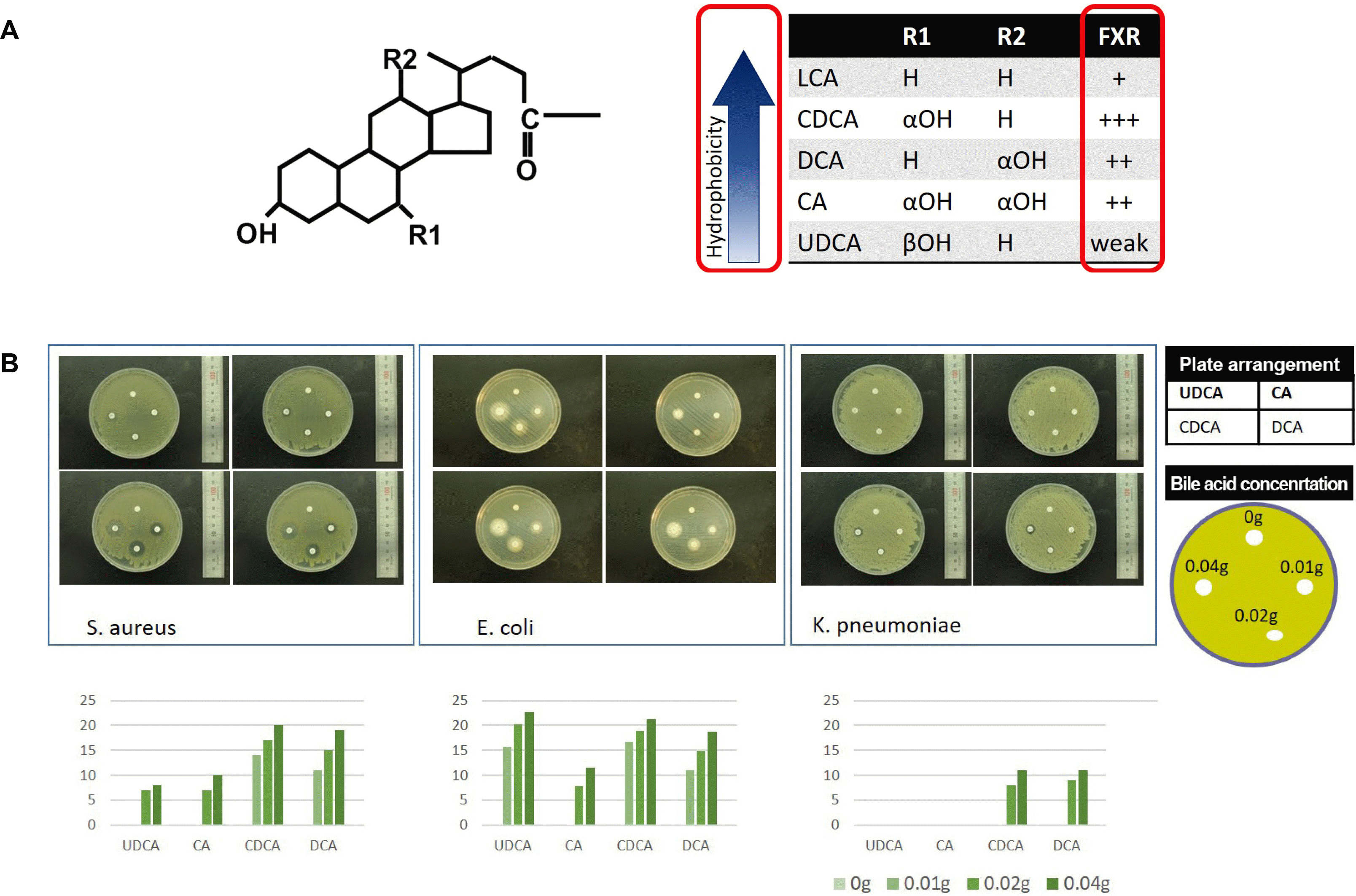 | Fig. 2Characteristics of bile acids in the body and various inhibitory powers against microbiotas. (A) The structure and characteristics of bile acids. (B) Bile acid susceptibility tests using the bile acids disk-diffusion methods. FXR, farnesoid X receptor; LCA, lithocholic acid; CDCA, chenodeoxycholic acid; DCA, deoxycholic acid; CA, cholic acid; UDCA, ursodeoxycholic acid; S. aureus, Staphylococcus aureus; E. coli, Escherichia coli; K. pneumoniae, Klebsiella pneumoniae.
|
담도계를 통하여 장관 내로 배출된 담즙산은 고유의 역할을 하면서 소장 내에서 대부분(약 95%) 재흡수되어 장-간순환을 통하여 재활용된다. 남은 약 5% 정도의 소량만 대변을 통하여 배출되며, 간에서는 매일 체외로 배출된 약 5% 정도 (500-600 mg)의 담즙산이 재생산된다(Fig. 3).58 각 개인의 종류별 담즙산 양과 조성도 이러한 과정을 통하여 일정하게 유지된다. 만약 특정 담즙산을 경구로 지속 공급하게 되면 공급하는 담즙산의 일부는 지속적으로 문맥계로 흡수되어 간으로 이동하게 되며 연속적인 장-간순환으로 전체 담즙에서 차지하는 비율이 점점 상승하여 주요 담즙산이 된다. 담즙산제 치료 요법은 현재 간담도계 질환에 대중적으로 사용되고 있으며, 그 대표적인 것이 우루소데옥시콜산(ursodeoxycholic acid, Urusa®; Daewoong Pharmaceutical Co.,Ltd., Seoul)이다. 우루소데옥시콜산은 케노데옥시콜산의 입체이성체(epimer)이며 정상적으로 인체 내에 5% 미만으로 존재하나 경구로 지속 공급하면 체내에서 총 담즙산의 50% 정도까지 상승하게 된다. 이외에도 과거 케노데옥시콜산이 담석 용해요법에 사용되기도 하였으며, 최근에는 obeticholic acid (Ocaliva®; Intercept Pharmaceuticals, Inc., New York City, NY, USA)와 같은 반(半)합성(semi-synthetic) 담즙산들도 개발되어 원발성 담즙성 담관염(primary biliary cholangitis)에 대한 치료 요법으로 사용되고 있다.59 담즙산 조성의 변화는 대중적으로 이용되는 담즙산 경구 복용으로 달성될 수 있으며, 결과적으로 장내 미생물 환경에 영향을 줄 것으로 추정된다.
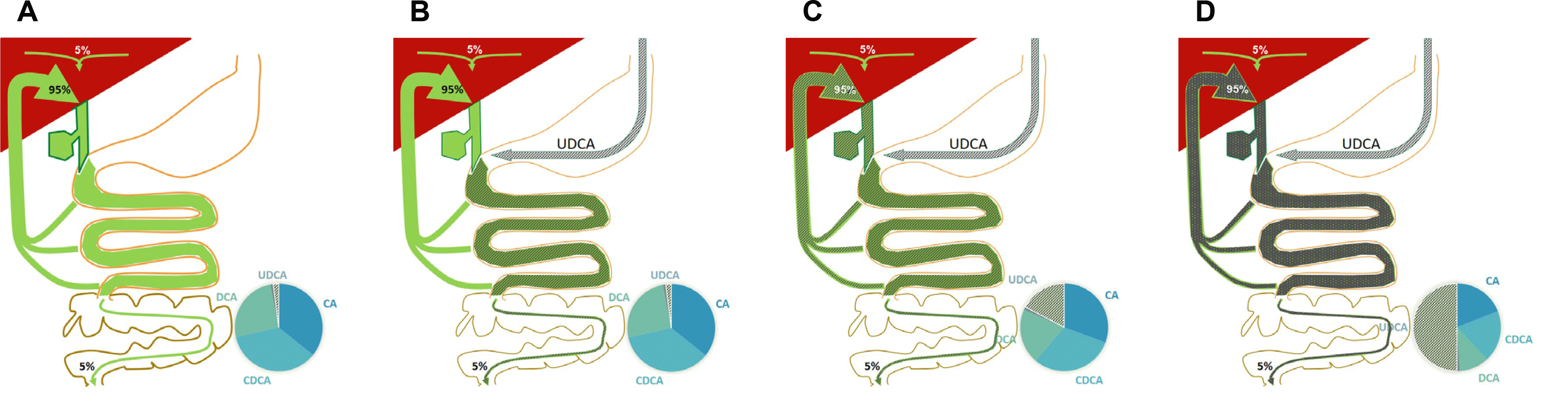
췌장으로부터 분비되는 외분비액에는 다량의 지질 및 단백분해효소 이외에 위장관의 미생물에 대한 내인성 면역에 관여하는 항균 펩타이드가 포함되어 있다. 항균 펩타이드는 주로 장관 내 파네트세포(paneth cell)에서 분비되나 췌장에서도 분비되며 췌장액 단백의 10%를 차지한다.60 항균 펩타이드는 선방세포(acinar cell; defensin α1)나 도세포(islet cell; defensin β) 혹은 둘 모두(defensins β2 and α4, cathelicidinrelated AMP [CRAMP])에서 분비된다.61-63 이는 장내 미생물과 상호작용을 하는데 췌장 내 면역계를 통하여 이루어진다. 예를 들어 β세포의 CRAMP 생산은 장내 미생물이 분비하는 단쇄지방산(short chain fatty acid)을 매개로 하여 췌장 내 면역 계통을 조절한다.62 CRAMP 증가는 tumor necrosis factor 생산을 감소시키고 tumor growth factor-β를 증가시키는 기전을 통하여 췌장 내 대식세포를 염증형(inflammatory phenotype)으로부터 조절형(regulatory phenotype)으로 변화시키고 추가적으로 조절 T 세포(regulatory T cells)와 기존의 수지상세포(conventional dendritic cells)를 유도하여 면역 조절 환경을 구축한다.61 결국 항균 펩타이드와 단쇄지방산과 같은 매개 물질에 의하여 형성된 양방향 신호전달 체계는 미생물의 수와 조성을 일정하게 유지시켜준다. 췌장의 외분비는 저장-작동식 칼슘통로(store-operated calcium channel)와 관련이 되어 있는데,64 Ahuja 등62은 그 기능을 저하시킨 후 장내 미생물의 과증식으로 인한 쥐의 사망을 보고하였다.
각 개체들은 장관 내에 특징적인 미생물 분포 및 구성을 가지고 있으며, 이는 각자의 특징적인 담즙산과 항균 펩타이드를 포함한 내인성 항생 물질 구성과 관련이 있을 것이라고 생각된다.65 이 때문에 외부에서 특정한 미생물이 장관 내로 공급되어도 대부분이 일정 시간이 지나면 내인성 항생 물질에 의하여 조절이 되어 원래의 장관 내 미생물 환경으로 회귀하게 될 것으로 추정된다. 췌담도계 질환은 이러한 내인성 항생물질의 생성 및 흐름에 영향을 줄 것으로 생각되며, 국내에서도 장내 미생물과의 관계에 대한 연구들이 진행되었다. 급성 및 만성 췌장염 환자에서 호기 수소(H2)가스와 메탄(CH4)가스 농도 측정상 소장 내 미생물의 과증식이 추정됨을 보고하였고, 이는 담석 질환자와 담낭 절제 후 환자에서도 관찰되었다.66-69 이 연구들에서는 측정 결과와 증상과의 관계를 함께 분석하여 췌담도 질환에서 관찰되는 복부 증상이 장내 미생물 변화에 의한 기능성 증상일 가능성을 제시하였다.
3. 췌담도 질환과 장내 미생물
담도계 질환은 미생물 감염과 함께 염증성 질환을 동반하고 있는 경우가 많고 대부분이 담즙의 흐름 장애와 정체 및 담도 폐쇄에 의하여 발생하는 경우이다. 해부학적 독립 구조, 담즙의 일방향 흐름, 담즙산의 미생물 억제 능력에도 담도계는 무균 환경이 아니며 많은 연구자들은 균의 기원이 장관 내 미생물인 것으로 추정하고 있다.5,6 실제 Salmonella typhi는 담즙산에 억제되지 않고 담도계에 잠복 감염 상태로 유지되기도 한다. 장내 미생물의 이동은 장관 내로부터의 역류와 혈액 및 림프계통을 통하여 이루어질 것으로 추정하고 있다.70 앞서 설명된 바와 같이 담즙산 조성의 개인차는 감염균에도 영향을 줄 것으로 생각된다.
최근에는 담도계 미생물이 악성 종양이나 담도 결석 등의 질환과 관계가 있음을 밝히는 문헌들이 보고되고 있다.6,9-13 질환자들의 담즙에서 확인된 미생물 환경이 질환의 원인인지 질환으로부터 영향을 받은 것인지 분명하지는 않다.10 그러나 Chen 등9이 보고한 결과에 따르면 담도 감염을 발생시킨 담도계 해부학적 변화의 원인이 악성 종양에 의한 경우와 결석에 의한 경우 사이에 검출된 균종의 명확한 차이가 있어 장내 미생물 환경의 변화가 질환의 발생 과정과 연관이 있을 가능성을 제시하였다. 이외에도 많은 연구들에서 보고된 담도계 감염균주들은 대부분 장내 미생물이었으며, 장내 미생물 환경이 담도계 질환과 무관하지 않음을 알 수 있다. 이러한 미생물들이 담즙 내에서 저항할 수 있는 것은 여러 기전이 있으며 대표적으로 multidrug resistance (MDR) 배출펌프에 의한 것이고,48,71-74 bile salt hydrolase (BSH) 활성도에 의해서도 영향을 받는 것으로 알려져 있다.75 췌장은 강력한 지방, 단백분해효소와 함께 항균 펩타이드를 생산, 분비하는 기관임에도 조직 내에서 미생물이 검출된다.76-78 기전은 담도계와 마찬가지로 장내 미생물이 췌관으로 역류, 혈액 및 림프계통을 통한 이동 등에 의할 것으로 생각된다.7,78-80 특히 췌장염의 경우 장내 미생물의 이동은 claudin 4의 감소에 의한 장관의 투과력 증가로 인하여 발생하며, 환자의 예후와 큰 연관이 있다.80-83 급성 췌장염의 경우 비질환자에 비하여 박테리오데테스와 프테오박테리아가 증가하고 피르미쿠테스와 아시네토균이 감소하는 것으로 알려져 있다.
자가면역췌장염의 환자에서도 장내 미생물의 변화가 있음이 보고되었으며,8,84,85 그중 Hamada 등85은 자가면역 이외의 원인에 의한 만성 췌장염 환자와 비교하여 명확한 차이가 있음을 확인하였다. 당뇨에 있어서도 1형 당뇨는 면역학적 매개에 의한 췌장의 β세포의 파괴가 주된 기전이며 인슐린 저항성이 주된 기전인 2형 당뇨와 구분된다. de Goffau 등86,87은 당뇨가 발생하기 전 1-5세 영아들을 대상으로 1형 당뇨 자가면역항체가 2개 이상 있는 아이와 그렇지 않은 아이들의 장내 미생물을 비교하였고, 젖산 생산형(lactate-producing) 박테리아와 낙산염 생산형(butyrate-producing) 박테리아의 수가 정상 아이들에 비하여 감소되어 있음을 확인하였다. Vatanen 등88은 1형 당뇨 가족력이 있는 신생아를 분석한 결과 단쇄지방산을 생산하는 박테리아가 감소되어 있음을 보고하여 단쇄지방산의 1형 당뇨 발생에 대한 보호 효과 가능성을 제시하였다. 췌장암에 대한 연구들도 진행되었는데, Ren 등89은 췌장암 환자에서 박테리오이데테스가 증가하였으며 정상 대조군에서는 피르미쿠테스와 프로테오박테리아가 증가한 것을 보고하였다.
4. 장관 내 미생물 환경 조절 요법
미생물의 경구 섭취 요법은 현재 가장 대중적으로 이용되는 방법이며, 프로바이오틱스(probiotics)나 미생물 촉진인자인 프리바이오틱스(prebiotics)를 경구로 투여하는 방법이다. 특정 균주를 강한 산성 환경의 위장(stomach)과 많은 분해 효소(pancreatic enzyme)가 존재하는 십이지장을 거쳐 작용 부위인 소장 및 대장에 도달하게 해야 한다. 이 때문에 균주들이 살아서 도달할 수 있도록 많은 전달체들이 개발되고 있으나 앞에서 기술한 바와 같이 소장에 도달한다 하더라도 미생물을 억제하는 다량의 담즙산이 존재하고 있어 결국은 내인적 조절자인 담즙산 조성에 영향을 받게 될 수밖에 없다. 다량의 균주를 전달하여 일시적인 수의 증가가 있을 수 있으나 결국 원래의 상태로 돌아올 가능성이 높고 지속적인 공급이 이루어져야 한다. 항생제 투여법은 장관 내 균주에 작용하여 증식을 억제하는 방법이며 목표 균주에 따라 다양한 종류의 정맥 및 경구 약물들이 있다. 단점은 일정 시간 이상 투여 시 미생물의 항생제에 대한 저항성이 발생하여 효율이 감소하게 되며, 단기간 요법으로 사용되게 된다. 대변 이식(fecal transplantation) 요법은 장내 세균 불균형으로 인하여 발생한 질환에 대하여 특별한 질환이 없는 정상인의 대변을 장관 내로 전달함으로써 병원균의 증식을 억제하는 방법이다. 현재는 Clostridium difficile에 의하여 유발되는 위막성 대장염(pseudomembranous colitis)에 대하여 치료 요법으로 사용되고 있고, 염증성 장질환, 과민성 장증후군, 비만, 당뇨 등 다른 질환에 대한 치료법으로도 시도되고 있다. 그러나 불특정 미생물을 다량 공급하여 다른 질환을 발생시킬 수 있고, 대상자의 면역계에 미치는 영향에 대하여도 장기적인 데이터가 부족하다. 췌담도계에서 배출된 담즙산이나 췌장의 외분비액은 장관 내 미생물 환경을 조절하는 주 조절자이며 해부학적 변화(결석, 종양)에 대한 물리적 치료나 합성 담즙산 공급 등의 약물 요법에 의하여 제한된 범위에서 장내 미생물 환경에 영향을 미칠 수 있다. 특히 대중적으로 사용되고 있는 경구담즙산 공급 요법은 목표 미생물을 조절하는 방법이 될 수 있다.
Go to :
결 론
장내 미생물(gut microbiota)은 인체의 일부로서 신체 대사에 관여하고 다양한 질환의 발생과 연관이 있다. 장관 내에 서 이들의 유전 정보(gut microbiome)를 검출하여 분석하는 것은 인간의 유전자(human genome)를 분석하는 것만큼 중요하다. 장내 미생물에 대한 유전 정보 중 모든 인간이 공유하고 있는 주요 기능성 유전자(core microbiome)들은 인체의 생리적 기능을 이해하는 데 도움을 주고 질환자의 장내 미생물 유전 정보는 질환의 진단 및 치료에 이용될 수 있다. 췌담도계는 장내 미생물 환경에 직접적인 영향을 미치는 항균 기능 물질들(담즙산, 항균 펩타이드)을 장관 내로 배출하며, 장내 미생물은 배출된 물질들의 조성을 변화시키거나 간접적인 신호 전달을 통하여 항균 기능 물질의 생성 및 배출을 조절하는 방법으로 췌담도계에 영향력을 미친다. 이러한 양방향 소통(crosstalk)은 췌담도계 분비 물질과 미생물의 항상성을 유지시키며 이것이 제대로 작동하지 못하면 미생물 불균형(dysbiosis)과 질환을 발생시키게 된다. 담즙산제 치료 요법은 기존에 대중적으로 시행되고 있는 방법으로 장내 미생물 환경에 영향을 준다. 악성 종양을 포함한 많은 췌담도 질환에서 장내 미생물의 연관성이 보고되고 있다. 전통적으로도 대부분의 췌담도 질환은 장내 미생물의 감염이 동반되어 있어 치료의 중요한 대상이 된다. 췌담도계는 물리적, 약물적 치료 요법을 통하여 그들의 기능을 조절할 수 있다. 이는 장내 미생물에 대한 연구나 치료에 있어 새로운 개척 분야가 될 수 있다.
Go to :
 XML Download
XML Download