

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Alcoholic liver disease (ALD) is a common chronic liver disease worldwide that is caused by the overconsumption of alcohol. This could cause steatosis, steatohepatitis, fibrosis, and liver cirrhosis. The development of ALD is complex and has been linked to oxidative stress, including changes in the redox state, oxidative metabolites of ethanol, and effects on antioxidants [1]. For example, ethanol induces the enzyme, CYP2E1 (P4502E1), an ethanol oxidizer, to generate free radicals that cause liver injury [2]. Reactive oxygen species (ROS) inhibit AMP-activated protein kinase (AMPK) and increase the expression of sterol regulatory element-binding protein (SREBP-1), leading to increased cholesterol synthesis [3]. In addition, the ethanol metabolism via acetaldehyde could affect peroxisome proliferator-activated receptor (PPAR) α, which regulates the hepatic fatty acid metabolism, resulting in the accumulation of triglyceride (TG) in the liver [4]. Furthermore, ethanol inhibits the hepatic antioxidant, glutathione (GSH), which leads to mitochondrial toxicity by H2O2.
Many pathogeneses of ALD focused on a gut barrier dysfunction. Alcohol-induced endotoxin, an integral component of the Gram-negative bacterial outer membrane, activates Kupffer cells to secrete cytokines, and ultimately cause liver damage [567]. Therefore, altered intestinal microflora could be one of the therapeutic targets for ALD [89]. Lactobacillus rhamnosus GG was reported to reduce the endotoxemia and liver damage caused by alcohol [10]. Another report also showed that the consumption of L. rhamnosus GG for 2 weeks led to a decrease in endotoxemia and liver steatosis in C57BJ/6N mice with ALD [11]. A human clinical trial revealed a decrease in the aspartate transaminase (AST) and alanine transaminase (ALT) levels in the serum of ALD patients, who consumed Bifidobacterium bifidum and L. plantarum 8PA3 for 5 days [12]. The gut microbiota plays a vital role in controlling the development of ALD.
Probiotics can change the gut microbiota profile and improve the gut barrier [1314]. One of the widely studied probiotic bacteria, L. rhamnosus GG, helped prevent an alcohol-induced leaky gut and liver injury [151617]. On the other hand, few studies have examined the effects of other Lactobacillus strains on liver disease. Therefore, this study examined different lactic acid bacteria (LAB) species L. casei GKC1 (GKC1), L. fermentum GKF3 (GKF3), Bifidobacterium lactis GKK2 (GKK2), L. rhamnosus GKLC1 (GKLC1), L. paracasei GKS6 (GKS6), and L. plantarum GKM3 (GKM3) on alleviating the development of ALD. This study aims to provide a better understanding of probiotic strains as another option of a therapeutic approach for patients with chronic liver disease.
MATERIALS AND METHODS
Probiotic supplementation
The bacterial sources GKC1 (BCRC 910825), GKF3 (BCRC 910824), GKK2 (BCRC 910826), GKLC1 (BCRC 910828), GKS6 (BCRC 910788), and GKM3 (BCRC 910787) were obtained from Grape King Bio Ltd. (Taoyuan, Taiwan) were preserved at BCRC, Taiwan. The bacteria were cultured in MRS broth for 16 h at 37°C and centrifuged at 3,800 rpm under 20°C for 10 min. The supernatant was removed, and the bacteria pellet was collected. After mixing with 5% skimmed milk, the mixture was treated immediately with vacuum freeze-drying (Kingmech Scientific Co., Taipei, Taiwan). The total cell numbers in the LAB freeze-dried powder were measured by plate counting and Guava® easyCyte™ Flow Cytometers (Luminex Crop., Austin, TX, USA).
Animal subjects
The animal use protocol in these experiments was reviewed and approved by the Institutional Animal Care and Use Committee (No. 106-13f). Male C57BL/6 mice (8-week-old) weighing approximately 25 g were housed at 22 ± 3°C with a 12:12-h dark-light cycle. During the experiment, mice (n = 48) were divided into 8 groups as follows: 1) Normal control, mice fed with a normal liquid diet (PMI® Micro-Stabilized Rodent Liquid Diet LD101); 2) Positive control, mice fed with alcohol-containing liquid diet (PMI® Micro-Stabilized Alcohol Rodent Liquid Diet LD101A) (36% calories were provided from alcoholic liquid diet); and 3) Probiotics group, an alcohol-containing diet with additional probiotics GKS6; 4) GKM3; 5) GKLC1; 6) GKC1; 7) GKF3; and 8) GKK2. The feeding dose was 0.82 g/kg B.W. per day by oral gavage. The blood samples of the mice were collected every 2 weeks for biochemistry analysis. After 8 weeks of the experiment, the animals were sacrificed, and their liver tissues were collected for both biochemistry analysis and histology investigation.
Biochemistry analysis
At weeks 0, 2, 4, 6, and 8 (sacrificed), the sera of the mice after each treatment were collected by centrifuge a t 14,000 × g for 5 min at 4°C, and the AST, ALT, and glucose levels were analyzed (7070 Autoanalyzer; Hitachi, Tokyo, Japan).
GSH and catalase (CAT) analysis
Liver tissue (0.5 g) mixed with a ten-fold volume of liver homogenate (8 mM K2HPO4, 12 mM KH2PO4, 1.5% KCl, pH 7.4) was centrifuged at 10,000 × g for 15 min at 4°C. The supernatant was collected and measured using a commercial kit from Cayman Chemical Company for GSH (703002) and CAT (703102).
TG and cholesterol measurement
The liver (0.2 g) was ground with 4 g of Folch reagent (chloroform: methanol= 2:1, V/W) in a tissue homogenizer (HsiangTai, New Taipei, Taiwan) overnight [18]. The extract was filtered through a Whatman No. 1 paper and stored at 4°C. The liver extract, 0.2 mL and 0.5 mL were dried to measure the TG and total cholesterol (TC) levels, respectively. A commercial kit (RANDOX Lab. Ltd., UK) was used to measure the TG and TC levels from their absorption at 500 nm using a spectrophotometer (Epoch™; BioTek, Winooski, VT, USA).
Liver histology and liver fat
The liver tissues were fixed in 10% formalin and processed to 5 μL thick slices using a microtome (Leica RM 2145; Leica Microsystems, Nussloch, Germany). The samples were stained with haemotoxylin and eosin and examined using an optical microscope (Opticphot-2; Nikon, Tokyo, Japan). The histopathology was assessed by a veterinarian from BioLASCO Taiwan Co., Ltd. The liver fat was graded from 0 to 4, which represented the percentage of liver cells containing lipid 0%, < 10%, 10%–33%, 33%–66%, and 66%–100%, respectively, in the histology sections according to the standard based on Ryu's study and formulated by the Food and Drug Administration, Taiwan [19]. The histopathology of the stomach and gut were scored from 1 (< 10%) to 4 (80%–100%) according to Shackelford’s report in the Journal of Toxicologic Pathology [20].
Statistics
All data are presented as the mean ± standard deviation. An independent-samples t-test (SPSS 16.0; SPSS Inc., Chicago, IL, USA) was applied for ALT analysis as the first stage of selection. The other parameters were evaluated using a Duncan's multiple range test of 1-way analysis of variance (SPSS 16.0). A P-value < 0.05 was determined to be the level of significance compared to the positive control.
RESULT
Table 1 lists the cell numbers contained in LAB freeze-dried powder. All alcohol-fed mice showed lower body weights than the normal-diet mice (data not shown) on week 8. Probiotic supplementation in the alcohol-fed mice did not contribute to the bodyweight compared to the positive control. The relative liver weights showed a similar tendency (data was shown).
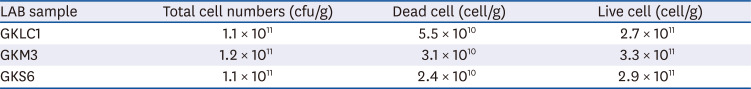
Table 1
Cell numbers in LAB freeze-dried power GKLC1, GKM3, and GKS6
| LAB sample | Total cell numbers (cfu/g) | Dead cell (cell/g) | Live cell (cell/g) |
|---|---|---|---|
| GKLC1 | 1.1 × 1011 | 5.5 × 1010 | 2.7 × 1011 |
| GKM3 | 1.2 × 1011 | 3.1 × 1010 | 3.3 × 1011 |
| GKS6 | 1.1 × 1011 | 2.4 × 1010 | 2.9 × 1011 |
LAB, lactic acid bacteria; cfu, colony-forming unit; GKLC1, Lactobacillus rhamnosus GKLC1; GKM3, Lactobacillus plantarum GKM3; GKS6, Lactobacillus paracasei GKS6.
![]()
AST, ALT, and glucose analysis
No significant differences in all the serum AST and ALT results were observed in the first week (P > 0.05). During the experiment, the ALT increased continuously in the positive control. In contrast, the serum ALT in probiotics GKS6, GKM3, and GKLC1 were only raised slightly (data not shown). After 8 weeks of feeding, the serum AST and ALT in the positive control were higher than the normal control (P < 0.05) (Fig. 1). On the other hand, the AST levels were reduced significantly in the GKLC1 group compared to a positive control (Fig. 1A). In the ALT analysis result, the mice fed GKS6 and GKM3 were significantly lower than the positive control (Fig. 1B). In addition, GKLC1 also showed a similar tendency (P = 0.052) on reducing the ALT levels. The expected GKK2 showed a decrease in AST. The other probiotic groups did not show significant differences (P > 0.05) in both AST and ALT when compared to a positive control (Supplementary Fig. 1). Therefore, the probiotics, GKS6, GKM3, and GKLC1, were selected from the other tested groups for further experiments. The concentrations of glucose in serum were increased from week 1 to week 8 in all experimental groups, and there were no significant differences observed between the groups (data not showed).
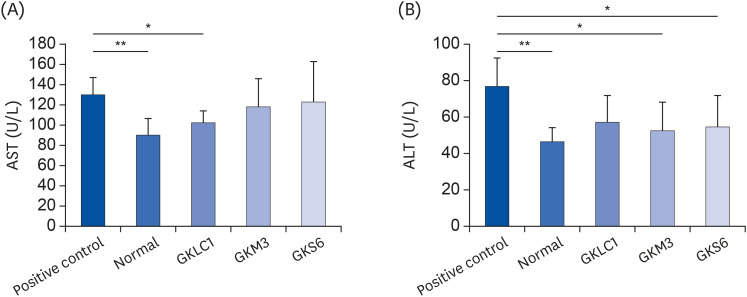
Fig. 1
Results of AST (A) and ALT (B) analysis at week 8. The results are expressed as mean ± SD. An unpaired t-test was applied (n = 6).
AST, aspartate aminotransferase; ALT, alanine aminotransferase; GKLC1, Lactobacillus rhamnosus GKLC1; GKM3, Lactobacillus plantarum GKM3; GKS6, Lactobacillus paracasei GKS6.
*P < 0.05, **P < 0.01 compared with positive control.
![]()
TG and TC
No significant differences in the serum TG and TC were observed among all the samples in the first week. At week 8, the serum TG after the GKM3 and GKLC1 treatment showed a significant reduction (P < 0.05) compared to a positive control (Fig. 2A); GKS6 showed a slightly decrease (P = 0.053). Similarly, the probiotic GKLC1 group showed a significant decrease in the levels of serum TC (P < 0.05) compared to the positive control on week 8 (Fig. 2B). The TG and TC concentrations in the liver of the non-alcohol fed mice were lower than in the positive control. Among alcohol-fed mice, the LAB treatments, GKS6, GKM3, and GKLC, produced significant decreases (P < 0.05) in both the liver TG and TC levels (Fig. 3).
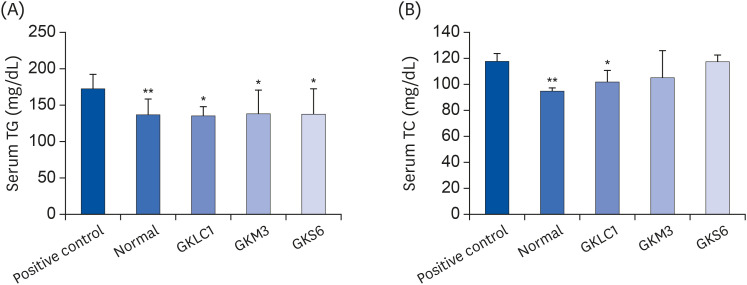

Fig. 2
Effect of probiotics GKLC1, GKM3, and GKS6 on the change in the serum TG (A) and TC (B) levels in ethanol-fed C57BL/6 mice at the 8th week. Each value is represented as the mean ± SD (n = 6).
TG, triglyceride; TC, total cholesterol; GKLC1, Lactobacillus rhamnosus GKLC1; GKS6, Lactobacillus paracasei GKS6; GKM3, Lactobacillus plantarum GKM3.
*P < 0.05, **P < 0.01 when compared to the positive control (Duncan's multiple range test).
![]()
Fig. 3
Effects of probiotics GKLC1, GKM3, and GKS6 on liver TG and TC in ethanol-fed C57BL/6 mice. Each value is represented as the mean ± SD (n = 6). Duncan's multiple range test was applied.
TG, triglyceride; TC, total cholesterol; GKS6, Lactobacillus paracasei GKS6; GKM3, Lactobacillus plantarum GKM3; GKLC1, Lactobacillus rhamnosus GKLC1.
*P < 0.05, †P < 0.01, ‡P < 0.05, §P < 0.01 compared to the positive control.
![]()
GSH levels and CAT activities
The GSH in the positive control was lower than the normal control (P < 0.05). On the other hand, the GSH activities in the selected groups were significantly higher than the positive control (P < 0.05) (Fig. 4A). The alcohol-fed mice without probiotics showed a lower CAT level than the non-alcohol fed mice. The GKS6 and GKM3 treatments resulted in a higher level of CAT compared to the positive control (P < 0.05) (Fig. 4B).

Fig. 4
Effect of probiotics GKLC1, GKM3, and GKS6 on the GSH (A) and CAT (B) activities in the liver of ethanol-fed C57BL/6 mice. Each value is represented as the mean ± SD (n = 6). Duncan's multiple range test was applied.
GSH, glutathione; CAT, catalase; GKLC1, Lactobacillus rhamnosus GKLC1; GKM3, Lactobacillus plantarum GKM3; GKS6, Lactobacillus paracasei GKS6.
*P < 0.05, **P < 0.01, ***P < 0.001 when compared to the positive control.
![]()
Histology
Fig. 5A shows the histopathology score in the liver, gut, and stomach. The histochemical stains showed that liver fat accumulation was occurred strongly in the alcohol-fed mice but decreased when treated with GKLC1, GKM3, and GKS6. (P < 0.05) (Fig. 5). Slight leakage was observed when treated with GKLC1, GKM3, and GKS6 in the gut, but strong necrosis was noted in the positive control (Fig. 5B). In the stomach histopathological score, less damage occurred in the GKLC1 group compared to the positive control (P < 0.05). In addition, less necrosis was observed in GKS6 and GKM3, but serious damage was noted in the alcohol-fed mice group without probiotics (Fig. 5B).
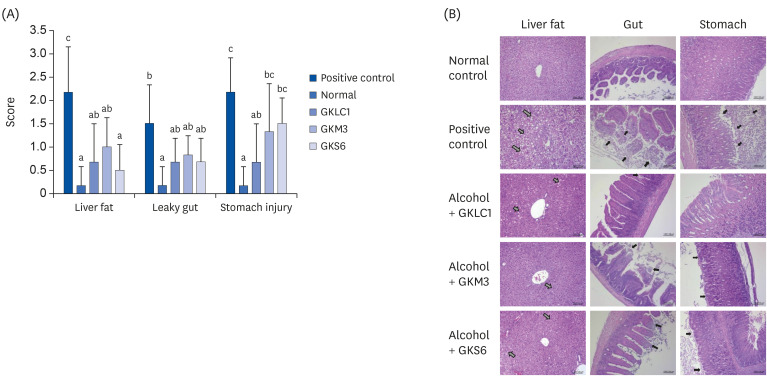
Fig. 5
(A) Histopathological score in the liver fat, leaky gut, and stomach of the ethanol-fed C57BL/6 mice with supplements of probiotics GKLC1, GKM3, and GKS6, respectively. The results are expressed as the mean ± SD. Different letters indicate a significant difference according to an analysis of variance (P < 0.05). (B) Histopathological analysis of mice in the liver, gut, and stomach of the mice tissue. The gray arrow indicates the accumulation of liver fat. The black arrow shows the location of the stomach or gut necrosis. GKLC1, Lactobacillus rhamnosus GKLC1; GKM3, Lactobacillus plantarum GKM3; GKS6, Lactobacillus paracasei GKS6.
![]()
DISCUSSION
Heavy alcohol consumption could develop ALD through the process of steatosis (fatty liver), hepatitis, fibrosis, and cirrhosis [21]. Generally, the accumulation of more than 5% TG as lipid droplets in hepatocytes is regarded as steatosis [22]. The histopathology result revealed the positive control to show a more than 10% difference in the fatty liver (histopathological score of level 2) than the normal control. This suggests that a hepatic lesion had been induced in mice by feeding them an alcohol liquid diet (Fig. 5). Because there was no significant change in body weight among the alcohol-fed mice, it was not expected that the excipient (skimmed milk) from the LAB powder would affect the calorie consumption significantly. Furthermore, there were no functional reports about milk on liver protection. Therefore, the liver fat and the liver TG and TC levels decreased significantly in the probiotics GKLC1, GKM3, and GKS6 groups suggests that these probiotics could prevent the process of steatosis (Fig. 3).
The earliest response of the liver to alcohol abuse is the accumulation of fat in hepatocytes [9]. Many studies have suggested that alcohol causes the accumulation of liver fat by gene regulation in both fatty acid synthesis and fatty acid oxidation [2324]. In a previous study on a high-fat diet (HFD) fed rats, higher levels of AMPK, PGC-1, and ACO mRNA expression, which are related to fatty acid oxidation, were observed in the livers of HFD-fed rats given supplementary of probiotics, GKM3, than in the other HFD-fed rats [25]. In addition, the levels of the PPAR-γ and SREBP-1c gene expression in the liver of HFD-fed rats were significantly lower in the GKM3 group, indicating the lower occurrence of fatty acid synthesis. The serum TC and TG in the alcohol-fed mice showed tended to reduce after 4 weeks of taking probiotics GKLC1, GKM3, and GKS6, respectively (data not shown). Therefore, the lower levels of liver TG and TC observed in alcohol-fed mice with probiotics groups (Fig. 3) were associated with the expression of these genes via the contribution from these probiotics.
Both AST and ALT are aminotransferases usually measured for a diagnosis of liver disease in clinical practice [26]. When the tissue is damaged, AST and ALT are released into the bloodstream. Therefore, increased aminotransferases levels in the serum indicate cell lysis. Both the serum ALT and AST levels could be reduced when the probiotics act on the gut barrier by reducing alcohol or endotoxin migration to the other organs and the phenomenon of cell lysis [262728]. Interestingly, the biochemical results in the serum AST and ALT showed a decrease in the GKLC1 groups, but only ALT reduction was detected in GKS6 and GKM3 compared to the positive control (Fig. 1). In alcohol hepatitis, AST is increased significantly more than ALT [29]. AST was found in a variety of tissues, including the liver, heart, skeletal muscle, kidneys, brain, and red blood cells, but ALT presents primarily in the liver. Therefore, an ALT measurement might interfere easily with liver injury [30]. The asynchronous of changes in the serum AST and ALT indicated that probiotics GKS6 and GKM3 might be associated with derivatives that play a role in protecting the liver protection. The biomarker, such as TNF-α, IL-6, INF-γ, and NF-κB, should be tested for further investigation. Note that the probiotics GKLC1 might function differently from GKS6 and GKM3 in alleviating ALD. The reduction of the liver index and the score of the histopathological sections in the GKLC1 group supported the liver and stomach protection (Fig. 5B).
Segawa et al. [31] reported that the heat-killed L. brevis SBC8803 could ameliorate ALD in mice. Probiotics might inhibit the endotoxins migrating into the liver through the improvement of the intestine barrier. Endotoxins are lipopolysaccharides derived from the cell walls of gram-negative bacteria. These endotoxins can migrate from the intestine to the liver when the epithelial barrier is disrupted by alcohol and cause liver damage by a proinflammatory process via an LBP/CD14/TLR-4-dependent mechanism [32]. Although the mechanisms of probiotics on preventing liver injury are not completely understood, it is generally thought that probiotics could protect against liver damage by improving the gut barrier [3334]. On the other hand, prepared probiotics contain live and dead cells, which might have different effects on liver protection. The mechanisms of dead cells on liver protection are believed to involve their cellular components [35]. Therefore, the samples were quantified by microflow cytometry to realize the dead/live cell ratio for judging their quality. The high percentage of live cells contained in the samples, which suggested that liver protection might occur mainly from the live cells (Table 1). In contrast to Segawa's study on the reduction of both serum ALT and AST, it was surmised that the probiotics not only alleviated ALD by improving the gut barrier, but their derivatives could also contribute to liver protection.
During ethanol oxidation, the alcohol-metabolizing enzyme, CYP2E1, catalyzes the electron oxidation of ethanol to acetaldehyde and involves free radical generation, which leads to oxidant stress and mitochondrial dysfunction [36]. The endogenous antioxidant, GSH, plays an important role in scavenging free radicals and is involved in regulating redox homeostasis [530]. Therefore, the higher GSH levels after the GKLC1, GKM3, and GKS6 treatments (Fig. 4A) suggest that these probiotics could prevent liver injury caused by ROS. The CAT present inside the liver mitochondria is also involved in oxidative stress defense by removing H2O2 to protect liver damage [37]. The restored CAT activities (Fig. 4B) confirmed that GKS6 and GKM3 could also affect the alcohol metabolism by regulating CAT in the peroxisomes. These results suggest that LAB GKLC1, GKM3, and GKS6 could reduce oxidative stress induced by alcohol and achieve the protective effects of developing ALD.
This study examined the effects of LAB GKC1, GKF3, GKK2, GKLC1, GKS6 and GKM3 on the protection of developing ALD. Three strains were selected, and it was demonstrated that the consumption of LAB powder GKLC1, GKM3, and GKS6 could alleviate ALD. This study had some limitations, such as lack of microbiota analysis to determine the probiotic interaction and the partial unclear mechanism for the pathway of preventing liver injury. Nevertheless, these results still show that probiotics could alleviate ALD development by modulating the lipid metabolisms, redox reaction, and gut barrier protection. The intake of GKLC1, GKM3, and GKS6 reduced the TG and TC concentrations in the liver, accumulation of liver fat, and the increase in GSH levels. The GKS6 and GKM3 treatment could restore the concentration of CAT affected by alcohol and the decrease in serum ALT. Overall, probiotics GKLC1, GKM3, and GKS6 could be promising agents or complementary therapeutic approaches in the prevention of ALD.
 XML Download
XML Download