

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Brain aging is a worldwide concern since it has been identified as the most critical risk factor for severe neurodegenerative diseases, including Alzheimer's disease and Parkinson's disease [1]. As a person ages, cells in the brain experience increased oxidative stress and metabolic impairment which can result in protein aggregation and DNA damage. These changes can subsequently manifest as increased anxiety and impaired cognitive function. In addition, the rate of neurogenesis decreases with age [2]. Both neuronal differentiation/neurite outgrowth and proliferation are important processes for neurogenesis [34]. Neurogenesis can be identified by analyzing β-tubulin III, a neuronal cell marker, and neurofilament heavy protein (NEFH), a major component of the neuronal cytoskeleton [5].
Aged brain tissue is characterized by down-regulation of major anti-oxidant enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GPX) [6]. It has been shown that plaques composed of aggregated amyloid β (Aβ) are leading causes of Alzheimer's disease pathology. Aβ is produced following cleavage of amyloid protein precursor (APP) by pro-apoptotic enzymes, such as caspases [7]. Brain aging has also been directly associated with neurodegenerative diseases characterized by increased chronic neuro-inflammation [8]. Correspondingly, higher levels of interleukin (IL)-8, IL-1β, and tumor necrosis factor (TNF)-α have been detected in aged animal brains [910].
D-galactose is a reducing sugar which converts to aldose and hydrogen peroxide when galactose oxidase presents at high concentrations. These by-products increase oxidative stress and accelerate the aging process [11]. A continuous low dose administration of D-galactose in mice has shown to induce changes which resemble accelerated brain aging [1213]. In particular, aging of various types of brain cells, including astrocytes [14], neuronal cells [15], and neuroblastoma cells [1617] have been observed in these models.
L-histidine is a common amino acid which is conditionally essential [18] and is a strong scavenger of hydroxyl radicals and singlet oxygen [1920]. In a previous study, supplementation with 1-10 mM L-histidine extended the lifespan of Caenorhabditis elegans [21]. L-histidine has shown to protect against diseases related to brain aging such as brain infarction [22], cerebral ischemia [23], brain edema [24], sciatic nerve lesions, and neuropathic pain [25]. In rats with acute liver failure, administration of L-histidine increased the anti-oxidant capacities of the brain cerebral cortex [26]. Moreover, L-histidine has shown to significantly attenuate ammonia-induced neurotoxicity by blocking glutamine transport into mitochondria in primary cultures of astrocyte [27].
L-carnosine is a natural dipeptide composed of β-alanine and L-histidine and it is present in human muscle and brain tissues [28]. Studies in male Drosophila melanogaster flies and senescence-accelerated mice have shown that L-carnosine supplementation can efficiently increase lifespan [2930]. Previous studies have also demonstrated that L-carnosine could exert neuroprotective effects against toxicity induced by N-methyl-D-aspartate, zinc, rotenone, and Aβ [31323334]. Moreover, L-carnosine attenuated early brain injuries by regulating oxidative stress and apoptosis in rats [35] and it was able to rescue cognitive decline induced by a high-fat diet in AD mice [36].
It has been demonstrated in previous reports that L-carnosine and L-histidine exert neuroprotective effects by activating the carnosine-histidine-histamine pathway [3133]. Moreover, carnosine is a dipeptide molecule which contains histidine [37]. However, while their close connection in terms of brain function and physiology is well-established, the neuroprotective effects of L-histidine and L-carnosine in combination remain poorly understood.
Despite these insights into the function of L-histidine and L-carnosine, their potential to mediate anti-aging effects in D-galactose-induced brain aging models has not been characterized. Therefore, in the present study, D-galactose-induced aging in retinoic acid (RA)-induced neuronal differentiated SH-SY5Y cells was investigated following exposure to L-histidine, L-carnosine, and a combination of both. In particular, brain neuronal regeneration, Aβ (1-42) production, apoptosis, antioxidation, and inflammation were examined.
Go to : 
MATERIALS AND METHODS
Cell culture and reagents
A human neuroblastoma cell line, SH-SY5Y, was purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). SH-SY5Y cells were maintained in a 1:1 mixture of Minimum Essential Medium and Ham's F-12 (Welgene, Daegu, Korea) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and 1% penicillin (100 U/mL)/streptomycin (100 µg/mL) (Invitrogen, Carlsbad, CA, USA). Cells were incubated in 5% CO2 at 37°C.
All-trans-retinoic acid (RA), D-galactose, L-histidine, and L-carnosine were purchased from Sigma-Aldrich (St. Louis, MO, USA). SH-SY5Y cells were treated with 10 µM RA for 6 d to differentiate them into neuronal cells. Stock preparations and treatments of D-galactose were performed under dim light due to light sensitivity of this sugar.
MTT assay
Neuronal proliferation was evaluated in MTT assays as described previously [38]. Briefly, differentiated SH-SY5Y cells were seeded onto 96-well plates (4 × 104 cells/well). Twenty-four hours later, the cells were treated with 200 mM D-galactose, 1 mM L-histidine, and 10 mM L-carnosine both individually, and in combination in media without FBS. After 48 h, media was removed and then cells were media containing MTT (500 µg/mL). After 4 h in 5% CO2 at 37°C, the supernatant of each well was discarded. Remaining formazan crystals were dissolved in 100 µL DMSO and absorbance values at 560 nm were recorded with a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Measurement of neurite length
Differentiated SH-SY5Y cells were treated with 200 mM D-galactose, 1 mM L-histidine and 10 mM L-carnosine individually and in combination in media without FBS for 48 h. Digital images of randomly chosen fields of cells were obtained with a light microscope (Olympus, Tokyo, Japan). Neurite length was measured with ImageJ software (National Institute of Health, Bethesda, MD, USA).
Assessment of cellular senescence
A commercially available senescence-associated β-galactosidase (SA-β-gal) assay kit was purchased from Cell Signaling Technology (Danvers, MA, USA) to examine cellular senescence. Briefly, differentiated SH-SY5Y cells were plated onto 6-well plates. Twenty-four hours later, cells were treated with 200 mM D-galactose, 1 mM L-histidine, and 10 mM L-carnosine individually, and in combination without FBS. After 48 h, the cells were rinsed with phosphate buffered saline (PBS) and then incubated with a fixative solution for 15 min at room temperature. After the cells were rinsed twice with PBS, they were incubated with β-gal staining solution at 37°C in the absence of CO2 overnight. Cells were imaged at 100x magnification with a light microscope (Olympus). Stained cells were counted in five randomly chosen fields at 100x magnification. The percentage of SA-β-gal-positive cells was calculated as follows: number of blue-stained cells / total number of cells.
Western blotting
Differentiated SH-SY5Y cells were treated with 200 mM D-galactose, 1 mM L-histidine, and 10 mM L-carnosine individually and in combination in the absence of FBS. After 48 h, protein concentrations were measured with a Bio-Rad Protein Assay Kit (Bio-Rad, Hercules, CA, USA). Briefly, protein samples were denatured, subjected to SDS-PAGE electrophoresis, and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). The, membranes were subsequently blocked with 5% bovine serum albumin or non-fat skim milk dissolved in Tris-buffered saline containing Tween 20 (TBS-T) at room temperature. After 1 h, the membranes were incubated with primary antibodies recognizing β-tubulin III (Sigma-Aldrich), NEFH (Sigma-Aldrich), APP (Abcam, Cambridge, UK), Aβ (1-42) (Cell Signaling), GPX-1 (Abfrontier, Seoul, Korea), SOD-1 (Santa Cruz Biotechnology, Dallas, TX, USA), cleaved caspase-3 (Santa Cruz Biotechnology), and β-actin (Abcam) at 4°C overnight. After the membranes were rinsed with TBS-T, they were incubated with secondary antibodies at room temperature for 1 h. To detect bound antibodies, an enhanced chemiluminescence reagent (Animal Genetics Inc., Suwon, Korea) was used and signals were detected on X-ray film.
Reverse transcriptase polymerase chain reaction (RT-PCR)
Differentiated SH-SY5Y cells were treated with 200 mM D-galactose, 1 mM L-histidine and 10 mM L-carnosine individually and in combination in the absence of FBS. After 48 h, RNA was isolated with Trizol reagent (Invitrogen) in accordance with the manufacturer's protocol. Quality confirmed RNA was used to produce cDNA by reverse-transcription with a Revert Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA). The cDNA generated was amplified with Taq polymerase (TAKARA, Tokyo, Japan) and primers specific for IL-8, IL-1β, TNF-α, and GAPDH as follows :(1) human IL-8: (forward) 5′-TGG CTC TCT TGG CAG CCT TC-3′ and (reverse) 5′-TGC ACC CAG TTT TCC TTG GG-3′ (2) human IL-1β: (forward) 5′-GGA TAT GGA GCA ACA AGT GG-3′ and (reverse) 5′-ATG TAC CAG TTG GGG AAC TG-3′ (3) human TNF-α: (forward) 5′-ACA AGC CTG TAG CCC ATG TT-3′ and (reverse) 5′-AAA GTA GAC CTG CCC AGA CT-3′ (4) human GAPDH (forward) 5′-AG AAG GCT GGG GCT CAT TTG-3′ and (reverse) 5′-AG GGG CCA TCC ACA GTC TTC-3′. PCR products were separated on a 2% agarose gel containing ethidium bromide and visualized under ultra-violet light. GAPDH was detected as an internal control.
Statistical analysis
All results are presented as the mean ± SEM of independent experiments performed at least three times. GraphPad Prism (GraphPad Software, Inc., La Jolla, CA, USA) was used to perform statistical analyses. Statistical significances between groups were determined by using one-way analysis of variance (ANOVA), followed by Newman-Keuls post-hoc test. P-values less than 0.05 were considered statistically significant.
Go to :
RESULTS
Effects of L-histidine, L-carnosine, and their combination on neuronal cell proliferation
To assess the effects of L-histidine, L-carnosine, and their combination on D-galactose-induced aged and RA-induced neuronal differentiated SH-SY5Y cell proliferation, MTT assays were performed. Cell proliferation was increased when the cells were treated with 1 mM (25.3%, P < 0.05) or 2 mM (29.0%, P < 0.05) L-histidine compared with the D-gal group (Fig. 1A). Similarly, L-carnosine treatment significantly increased cell proliferation at doses of 10 mM and 20 mM compared to the D-galgroup (33.0% and 42.1%, respectively; each P < 0.05) (Fig. 1B).
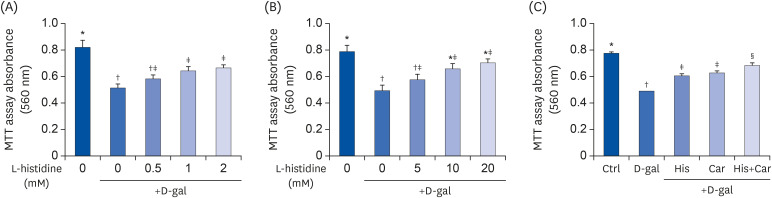 | Fig. 1Effects of L-histidine, L-carnosine, and their combination on proliferation.The neuronal proliferation was assessed by MTT assay. (A) L-histidine and (B) L-carnosine were treated with various concentrations for 48 h. (C) His, Car, and His+Car were treated for 48 h and assessed by MTT assay. The values shown are the mean ± standard error of the mean (n = 3–4).
Ctrl, Control; D-gal, 200 mM D-galactose; His, 1 mM L-histidine; Car, 10 mM L-carnosine; His+Car, 1 mM L-histidine + 10 mM L-carnosine.
*,†,‡,§Different superscript marks indicate significant differences between groups (P < 0.05).
|
We performed MTT assays to detect neuronal cell proliferation and to select suitable concentrations of L-histidine and L-carnosine for further experiments. In our preliminary experiments, differentiated SH-SY5Y cells were treated with L-histidine and L-carnosine at concentrations ranging from 0.5 mM to 50 mM for 48 h in the presence of D-galactose. Both L-histidine and L-carnosine improved neuronal proliferation in a dose-dependent manner. Statistically, significant effects on proliferation were observed between 1 mM and 10 mM, respectively. Thus, L-histidine exerted a 10-fold higher neuroprotective effect compared to L-carnosine in MTT assays.
Since our goal was to investigate effects of L-histidine and L-carnosine in combination against brain aging, we attempted to identify the minimum concentration exhibiting the similar neuroprotective effects. In addition, we performed MTT assays with various combination doses of L-histidine and L-carnosine. We analyzed neuronal cell proliferations in the presence of 1 mM L-histidine + 10 mM L-carnosine and in the presence of 2 mM L-histidine + 20 mM L-carnosine combinations. Both combination treatments exhibited significant synergistic effects compared with each treatment component alone. In contrast, 0.5 mM L-histidine + 5 mM L-carnosine in MTT assays did not show any additive effects. The combination of 1 mM L-histidine + 10 mM L-carnosine showed similar neuroprotective effects as the combination of 2 mM L-histidine + 20 mM L-carnosine (data not shown). Thus, these concentrations were used for L-histidine and L-carnosine in all subsequent experiments.
As shown in Fig. 1C, neuronal cell proliferation was significantly inhibited by D-galactose treatment compared with the Ctrl group (36.64%, P < 0.001). In contrast, L-histidine and L-carnosine treatments increased cell proliferation by 23.7% (P < 0.001) and 27.2% (P < 0.001), respectively, compared with the D-gal group. Furthermore, treatment with the combination of L-histidine and L-carnosine showed the largest increase in proliferation among the three groups (39.9%, P < 0.001).
Effects of L-histidine, L-carnosine, and their combination on neuronal cell regeneration
To characterize neuronal regeneration in RA-induced differentiated SH-SY5Y cells, average neurite length was measured. Compared with the Ctrl group, D-galactose treatment significantly decreased average neurite length. In contrast, treatment with L-histidine (40.80%, P < 0.01), L-carnosine (58.9%, P < 0.01), or both treatment group (76.23%, P < 0.001) significantly increased average neurite length compared with the D-gal group (Fig. 2A).
 | Fig. 2Effects of L-histidine, L-carnosine, and their combination on neuronal cell regenerations.Representative pictures for each group (100× magnification). (A) Ctrl, (B) D-gal, (C) His, (D) Car, and (E) His+Car. (F) The average length of the neurites of differentiated SH-SY5Y cells for each group was analyzed. ImageJ software was used to measure the individual neurite length. The protein expressions of an axonal marker, NEFH, and a neuronal marker, β-tubulin III were determined by Western blotting and β-actin was used as a loading control. (G) Representative blots are shown. Quantification of NEFH (H) and β-tubulin III (I) levels to β-actin are shown. The values shown are the mean ± standard error of the mean (n = 3–4).
NEFH, Neurofilament heavy polypeptide; Ctrl, Control; D-gal, 200 mM D-galactose; His, 1 mM L-histidine; Car, 10 mM L-carnosine; His+Car, 1 mM L-histidine + 10 mM L-carnosine.
*,†,‡,§Different superscript marks indicate significant differences between groups (P < 0.05).
|
NEFH is a major component of axonal cytoskeleton proteins [39] and β-tubulin III is a neuronal marker [40]. Thus, both of these proteins are widely used in neurobiology research. Here, NEFH and β-tubulin III were analyzed to investigate possible effects of L-histidine, L-carnosine, and their combination on neuronal regeneration (Fig. 2B). Expression levels of NEFH and β-tubulin III were 39.6% (P < 0.05) and 64.3% (P < 0.05) lower, respectively, in the D-gal group compared with the Ctrl group (Fig. 2C). In contrast, NEFH and β-tubulin III expression levels were higher following treatment with L-carnosine and following treatment with the L-carnosine and the combination than in the D-gal group. L-histidine also tended to increase the expression of NEFH and β-tubulin III, although not significantly in either case.
Effects of L-histidine, L-carnosine, and their combination on cellular senescence and levels of APP and Aβ (1-42) in neuronal cells
The SA-β-gal assay is a widely used method for verifying the presence of senescent cells [41]. We performed SA-β-gal staining to detect aging cells (Fig. 3A), the D-gal group exhibited 3-fold increase in the number of SA-β-gal positive cells compared to the Ctrl group (P < 0.001). Conversely, the numbers of SA-β-gal positive cells following treatment with L-histidine, L-carnosine, and a combination of both were significantly decreased compared with the D-gal group (His, P < 0.05; Car, P < 0.01; both, P < 0.01).
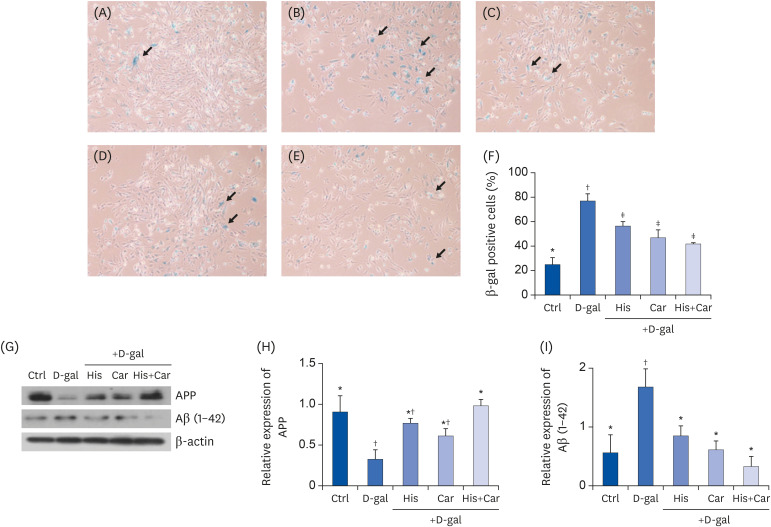 | Fig. 3Effects of L-histidine, L-carnosine, and their combination on cellular senescence and levels of APP and Aβ (1-42) in neuronal cells.SA-β-gal assay was performed to investigate cellular senescence. Representative images for each group at 100× magnification were shown. (A) Ctrl, (B) D-gal, (C) His, (D) Car (E) His+Car. (F) % of SA-β-gal positive cells in total counted cells. The protein expressions of APP and Aβ (1-42) were determined by Western blotting and β-actin was used as a loading control. (G) Representative blots are represented. Quantification of APP (H) and Aβ (1-42) (I) levels to β-actin are shown. The values shown are the mean ± standard error of the mean (n = 3–4).
APP, Amyloid β precursor protein; Aβ (1-42), Amyloid β (1-42); SA-β-gal, Senescence-associated β-galactosidase; Ctrl, Control; D-gal, 200 mM D-galactose; His, 1 mM L-histidine; Car, 10 mM L-carnosine; His+Car, 1 mM L-histidine + 10 mM L-carnosine.
*,†,‡Different superscript marks indicate significant differences between groups (P < 0.05).
|
Western blotting was also performed to determine whether treatment with L-histidine, L-carnosine, or their combination affected protein levels of APP and Aβ (1-42) in RA-differentiated SH-SY5Y cells (Fig. 3B). D-galactose treatment significantly down-regulated levels of APP (P < 0.05), whereas treatment with L-histidine and L-carnosine inhibited this decrease (P < 0.05). In addition, D-galactose treatment increased protein expression of Aβ (1-42) by 50% compared to the Ctrl group (P < 0.05). When cells were treated with L-histidine, L-carnosine, or their combination significant down-regulation of Aβ (1-42) expression by 50%, 64%, and 81%, respectively, was observed (His and Car, P < 0.05; His+Car, P < 0.01). Taken together, these results indicate that L-histidine and L-carnosine, when administered individually or in combination, they suppress D-galactose-induced aging-related Aβ formation.
Effects of L-histidine, L-carnosine, and their combination on expression of anti-oxidant enzymes and caspase-3 in neuronal cells
As a person ages, their brain experiences increased oxidative stress, metabolic impairment, and DNA damage [42]. In the present study, treatment with D-galactose decreased levels of the antioxidant enzymes, GPX-1 (P < 0.05) and SOD-1 (P < 0.05), and increased levels of the apoptosis marker, cleaved caspase-3 (P < 0.01) compared with the Ctrl group (Fig. 4A). In contrast, treatment with L-histidine, L-carnosine, or their combination recovered levels of expression of these proteins from D-galactose-induced decreases. For example, both L-carnosine and the combination of L-histidine and L-carnosine protected the level of GPX-1 which was down-regulated by D-galactose (P < 0.05 for both, respectively). Meanwhile, the level of cleaved caspase-3 was significantly down-regulated following treatment with L-histidine (37.2%, P < 0.05), L-carnosine (51.2%, P < 0.05), and their combination (54.6%, P < 0.05) compared to the D-gal group. Overall, these results suggest that L-histidine, L-carnosine, and their combination enhance anti-oxidation and inhibit apoptosis against D-galactose-induced aging in differentiated SH-SY5Y cells.
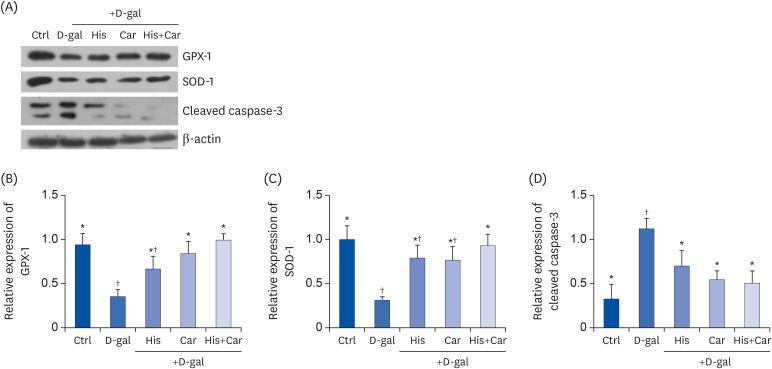 | Fig. 4Effects of L-histidine, L-carnosine, and their combination on anti-oxidant enzymes and caspase-3.The protein expressions of GPX-1, SOD-1, and cleaved caspase-3 were determined by Western blotting and β-actin was used as a loading control. (A) Representative blots are represented (left panels). Quantification of (B) GPX-1, (C) SOD-1, and (D) Cleaved caspase-3 levels to β-actin are shown. The values shown are the mean ± standard error of the mean (n = 3–4).
GPX-1, glutathione peroxidase-1; SOD-1, superoxide dismutase-1; Ctrl, Control; D-gal, 200 mM D-galactose; His, 1 mM L-histidine; Car, 10 mM L-carnosine; His+Car, 1 mM L-hisitidne + 10 mM L-carnosine.
*,†Different superscript marks indicate significant differences between groups (P < 0.05).
|
Effects of L-histidine, L-carnosine, and their combination on pro-inflammatory cytokines in neuronal cells
High-levels of chronic inflammation strongly contribute to the cognitive decline associated with brain aging. In particular, IL-8, IL-1β, and TNF-α are major inflammatory cytokines which are involved in the initiation and orchestration of neuro-inflammatory responses [4344]. To investigate the anti-inflammatory effects of L-histidine and L-carnosine both individually and in combination on D-galactose-induced inflammation, mRNA levels of IL-8, IL-1β, and TNF-α were analyzed in RA-differentiated SH-SY5Y cells (Fig. 5A). Following D-galactose treatment, mRNA levels of all three cytokines were up-regulated (IL-8 and IL-1β, P < 0.05; TNF-α, P < 0.001). In contrast, following treatment with L-histidine, L-carnosine, or their combination levels of IL-8, IL-1β, and TNF-α were restored to the levels observed in the Ctrl group.
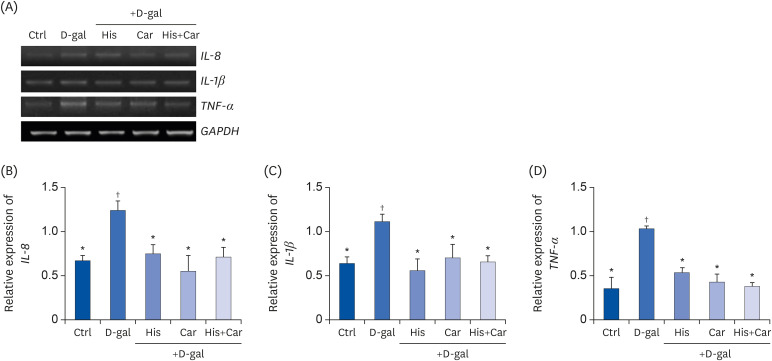 | Fig. 5Effects of L-histidine, L-carnosine, and their combination on pro-inflammatory cytokines in neuronal cells.The mRNA expressions of IL-8, IL-1β, and TNF-α were determined by RT-PCR and GAPDH was used as a loading control. (A) Representative blots are shown. Quantification of (B) IL-8, (C) IL-1β, and (D) TNF-α levels to GAPDH are shown. The values shown are the mean ± SE of the mean (n = 3–4).
Ctrl, Control; D-gal, 200 mM D-galactose; His, 1 mM L-histidine; Car, 10 mM L-carnosine; His+Car, 1 mM L-hisitidne + 10 mM L-carnosine.
*,†Different superscript marks indicate significant differences between groups (P < 0.05).
|
Go to :
DISCUSSION
In the present study, L-histidine, L-carnosine, and their combination exhibited anti-brain aging effects against D-galactose-induced aging in RA-differentiated SH-SY5Y cells. For example, these three treatments inhibited cellular senescence by restoring aging-related impairments. In addition, these treatments increased neurogenesis, neuronal proliferation, and expression levels of anti-oxidant enzyme, while also blocking Aβ formation, apoptosis, and proinflammatory mediators.
Both L-histidine and L-carnosine have been implicated as potent neuroprotective amino acids. Their ability to cross the blood brain barrier further supports this role [45]. L-histidine can penetrate blood brain barrier through a Na+-independent neutral amino acid transporter (system L) [46], whereas carnosine crosses blood brain barrier by passive diffusion. That's because its permeabilities are very low and similar to those of mannitol which has similar molecular weight with carnosine [4748]. Since L-histidine and L-carnosine do not compete each other when crossing the blood brain barrier, the combination treatments of L-histidine and L-carnosine would still enable themselves to be transported from blood to brain intact.
Daily consumption of histidine has shown to significantly improve working memory task capacity, clear thinking, and attentiveness [49]. Conversely, insufficient intake of L-histidine leads to diminished histamine levels in the brain. In mice, the latter is associated with anxiety-like behaviors [50]. Anti-brain aging effects of carnosine in a D-galactose-induced mouse model have also been reported [51]. For example, L-carnosine alleviated aging-induced increases in brain mitochondrial monoamine oxidase-A activity [52]. In addition, L-carnosine exerts neuroprotective effects via modulation of the HO-1/Hsp72 system and by reducing neuronal damage caused by oxidative stress [53].
The anti-brain aging properties of L-histidine and L-carnosine are closely related. This is consistent with L-carnosine being a component of L-histidine and β-alanine, thereby potentially providing similar physiological properties between L-histidine and L-carnosine [54]. Decker et al. [55] previously observed that carnosine was able to scavenge peroxyl radicals to a similar level as achieved by L-histidine. However, β-alanine did not exhibit the same effect. These results suggest that the radical scavenging capacity of carnosine is mainly dependent on L-histidine residue, especially its imidazole ring structure [5556]. Furthermore, considering that administration of L-carnosine increases the availability of L-histidine, neuroprotective activities of L-carnosine appear to be mediated by a carnosine-histidine-histamine signaling pathway in differentiated PC12 cells [31] and via rotenone-induced oxidative stress in brain microvascular endothelial cells [33]. L-histidine may be further converted into histamine, which has exhibited neuroprotective effects in both a neurodegenerative-induced hamster model [57] and a brain infarction model in rats [22]. Hiraga et al. [58] has suggested that L-histidine exerts its protective effects on ischemic events by facilitating the central histaminergic pathway which consequently inhibits recruitment of inflammatory cell by the histamine H2 receptor. However, conflicting reports have described that the neuroprotective activity of L-carnosine is independent of the histidine-histamine pathway, despite a partial role observed for the histamine H1 receptor [5960].
In the present study, D-galactose exposure significantly increased expression of Aβ (1-42) in differentiated SH-SY5Y cells. This effect was inhibited by treatment with L-histidine, L-carnosine, and their combination. L-carnosine disrupts hydrogen bond networks in the central hydrophobic cluster of Aβ (1-42). As a result, L-carnosine is able to inhibit aggregation of Aβ (1-42) [61]. In the present study, treatment with D-galactose increased Aβ formation. APP is a precursor of Aβ, and its APP cleavage can occur via amyloidogenic or non-amyloidogenic pathways. The former generates neurotoxic Aβ, whereas the latter blocks Aβ production and generates sAPPα, a molecule with neuroprotective functions [6263]. Based on these insights, we hypothesize that D-galactose up-regulates β-secretase and γ-secretase activities to generate Aβ from APP. Furthermore, these effects appear to be inhibited by L-histidine, L-carnosine, and their combination. Thus, additional studies are needed to measure and analyze levels of α, β, and γ secretases, as well as sAPPα, in order to confirm this hypothesis.
Consistent with the present results, previous in vivo studies have demonstrated that D-galactose induces down-regulation of anti-oxidant enzymes such as SOD and GPX [64], and up-regulation of the apoptosis marker, cleaved caspase-3 [65] in brain. L-histidine, L-carnosine, and their combination were found to efficiently restore expression of SOD and GPX, and to down-regulate the level of cleaved caspase-3 in the present study. L-histidine has exhibited anti-oxidant potential in brain and other tissues types by increasing the total anti-oxidant capacity of these tissues [66]. In addition, L-histidine has been found to increase the activity and mRNA level of GPX in the brain cerebral cortex in rat models of acute liver failure [26]. L-histidine has also been found to increase levels of mitochondrial glutathione (mGSH), a critical cell defense factor against oxidative stress, and GSH content in the prefrontal cortex of rats with acute hepatic encephalopathy [67]. It should be noted that the present results are also consistent with those of Aydin et al. [51]. The latter demonstrated that carnosine supplementation (250 mg/kg) significantly alleviates D-galactose-induced brain oxidative stress by regulating SOD and GPX activities, as well as the levels of cleaved caspase-3. In another study, carnosine (40 mg/kg) supplementation was found to improve GPX levels and oxygen radical anti-oxidant capacity in BALB/c mice receiving D-galactose [68].
A combination treatment of L-histidine and L-carnosine did not produce an additive effect, except in neuronal proliferation and neurite length analyses where the combination treatment induced a significantly better effect than L-histidine or L-carnosine alone. L-histidine and β-alanine which are precursors of carnosine, are able to cross the blood-brain barrier (BBB) [69] and undergo synthesis into carnosine in the forebrain, particularly in the olfactory bulbs [7071]. Despite the ability of carnosine to cross the BBB, most of the carnosine present in the brain originates from de novo synthesis rather than carnosine passing through the BBB [72]. Therefore, the combination treatment of L-histidine and L-carnosine may not produce a synergistic effect because their concentrations in the brain are tightly regulated through break down and synthesis reactions. Thus, their potentially additive effects may be inhibited. However, further studies are needed to investigate this hypothesis.
In conclusion, L-histidine and L-carnosine, both individually and combination, efficiently exerted protective effects in D-galactose-induced senescent neuronal cells in the present study. Specifically, these treatments improved neuronal proliferation, differentiation, anti-oxidation, apoptosis, and antiinflammation against D-galactose-induced neuronal impairment. These results support further studies of these amino acids as candidates for the treatment and prevention of age-related brain damage. In particular, the additional in vivo animal studies and clinical trials are necessary to confirm the neuroprotective effects of L-histidine, L-carnosine, and their combination in aged brains.
Go to :
 XML Download
XML Download