

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Hyperlipidemia is a common feature of dyslipidemia characterized by an increase of triglycerides (TG), total cholesterol (TC), and low-density lipoprotein cholesterol (LDL-C) and a decrease of high-density lipoprotein cholesterol (HDL-C) in the blood. The high levels of plasma lipid are strongly associated with a typical pathological condition of numerous diseases, such as atherosclerosis, non-alcoholic fatty liver diseases (NAFLD), and inflammation-related disorder [1234]. Dietary cholesterol is a well-known cause for hyperlipidemia [5]. Numerous in vivo studies demonstrated that a cholesterol-rich diet induces hyperlipidemia [5678].
Under the condition of hyperlipidemia, the inflammation and oxidative stress are predominant [9]. Dietary cholesterol-induced hyperlipidemia leads to an inflammatory response and enhances oxidative stress in organs [56]. In particular, hepatic inflammation plays a vital role in the progression of steatohepatitis, fibrosis, and finally, cirrhosis [10]. The excessive intake of cholesterol provokes hepatic inflammation, which directly results in the development of hepatitis [4]. Inflammatory responses are promoted by the release of inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1β, and IL-6, and inflammatory enzymes, such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), regulated by the nuclear factor-kappa B (NF-κB) activation [11]. In addition, elevated oxidative stress produces peroxynitrite and increases lipid peroxidation [12], which impairs the body's antioxidant status via downregulation of antioxidant enzymes, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GSH-Px) [13].
Chondroitin sulfate is a glycosaminoglycan, a type of polysaccharide that is present in the cartilages, skin, blood vessels, ligaments, and tendons of the body [14]. Chondroitin sulfate is mainly used for the treatment of osteoarthritis due to its anti-inflammatory activities [1415]. Besides, biological activities have established regarding the improvement of lipid/glucose metabolism, anti-atherosclerosis, antioxidant, and anti-apoptotic effects [16171819]. One of the major sources of chondroitin sulfate is shark cartilage. Recently, it has become necessary to replace shark cartilage-derived chondroitin sulfate (CS) because of the prohibition of the capture and killing of sharks [1920]. Therefore, several studies have made attempts to extract chondroitin sulfate from various sources, including cattle, pigs, chickens, and sea cucumbers [1921]. The skate (Raja Kenojei) is a commonly found fish species in the northwestern Pacific Ocean, Korea, Japan, China, and possibly elsewhere. Only the fillet of the skate is consumed due to its unique odor; the remaining parts (e.g. skin, cartilage, and bone) weighing about 30%, are discarded [22]. We previously investigated the protective effects of skate cartilage extracts containing chondroitin sulfate (SCS) in lipopolysaccharide (LPS)-induced liver damage [18]. However, it has not been studied whether SCS could alleviate hyperlipidemia-induced inflammation and oxidative stress in high cholesterol diet (HCD)-fed mice. Therefore, the aim of the present study is to investigate the beneficial effects of SCS on hyperlipidemia, inflammation, and oxidative stress in the LDL receptor knockout (LDLR-KO) mice fed an HCD in comparison with the effects of CS.
Go to : 
MATERIALS AND METHODS
Preparation of skate cartilage extracts containing chondroitin sulfate
The SCS was provided by Yeongsan Skate (Yeongsan Skate Co., Ltd., Busan, Korea). Skate cartilage was obtained during the skate processing as mentioned in the previous study [22]. Briefly, the byproduct of skate cartilage was heated at 95°C for 2 h to remove ribbon-type meat. Skate cartilage was dried for 24 h using a heated-air dryer and crushed. The powdered skate cartilage was added with 3 volumes of water and hydrolyzed by protease such as alcalase and protamex (Novozyme Co., Bagsværd, Denmark) at 50°C for 4 h, followed by heated at 95°C for 30 min. Afterward, it was deodorized, filtered, and concentrated at 65°C. The final sample was freeze-dried at −75°C. The composition of SCS was as follows; moisture 2.09%, protein 46.97%, ash 3.5%, and chondroitin sulfate 47.44% (Food Analysis Center, Pukyong National Univ., Busan, Korea). The positive control, CS was purchased from Sigma-Aldrich (C4384; Sigma-Aldrich, St. Louis, MO, USA).
Animal and experimental diets
Forty-five male LDLR-KO mice (6-week-old, each 20-25 g) were obtained from Jackson Laboratories (Bar Harbor, Me, USA). Mice were housed in a plastic cage with a 12 h light-dark cycle. The room had standard laboratory humidity (50 ± 10%) and temperature (20 ± 2°C). After 1 week of acclimatization, mice were randomly assigned into five groups on the basis of body weights (n = 8 per group). The experimental groups were shown in Table 1. The dose of CS and SCS was determined according to a previous report [18]. In particular, the concentration of SCS was considered by the content of chondroitin sulfate in SCS (47.44%) [18], thus the dose of SCS groups is a half of that of CS groups. CS and SCS were orally administered via a stomach tube every day for 10 consecutive weeks. During the experimental period, mice were provided with free access to water and HCD composed of 20 kcal% protein, 45 kcal% carbohydrate, 35 kcal% fat (D12336, Research Diets, New Brunswick, NJ, USA). The percentage of cholesterol in HCD was 1.25%. Body weight was recorded every week, and food intake was examined every day. The food efficiency ratio (%) was calculated as total body weight gain/total food intake × 100.
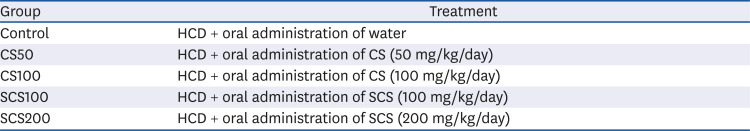
Table 1
The experimental groups in this study
![]()
The mouse was fasted for 12 hours before sacrifice, and anesthetized with CO2 gas. Immediately the blood was collected and centrifuged at 3,000 rpm for 20 min at 4°C to obtain serum. Liver tissues were perfused with 0.9% sodium chloride and removed. Serum and liver tissues were stored frozen in a deep freezer at −80°C. All experimental procedures were permitted (Approval No. PNU-2018-1887) using the guidelines established by the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC).
Serum lipid profiles, atherogenic index, and cardiovascular risk index
Serum triglyceride (TG, AM157S-K, Asan Pharm., Seoul, Korea), total cholesterol (TC, AM202-K, Asan Pharm, Seoul, Korea), and high-density lipoprotein cholesterol (HDL-C, AM203-K, Asan Pharm, Seoul, Korea) concentrations were measured using commercially available kits. Low-density lipoprotein cholesterol (LDL-C) concentration was calculated as TC - HDL-C / (TG / 5) [23]. The atherogenic index (AI) was calculated as (TC - HDL-C) / HDL-C, and the cardiac risk factor (CRF) was calculated as TC / HDL-C.
Serum inflammatory cytokine concentration
Serum inflammatory cytokine concentrations were measured using ELISA kits. The following cytokines were measured: TNF-α (#560478, BD, Franklin Lakes, NJ, USA), IL-1β (MLB00C, Minneapolis, MN, USA), IL-6 (#550950, BD, Franklin Lakes, NJ, USA), and prostaglandin E2 (PGE2, M6000B, Minneapolis, MN, USA).
Lipid peroxidation in the liver tissue
The content of malondialdehyde (MDA) in liver tissue was measured according to the method of Ohkawa et al. [24]. After centrifugation at 3,000 rpm for 10 min, the supernatant was mixed with 1% phosphoric acid and 0.67% thiobarbituric acid TBA), and the mixture was boiled for 30 min and then cooled. Seven milliliters of butanol was added and the mixture was centrifuged at 3,000 rpm for 10 min. The absorbance of the supernatant was measured at 540 nm. The standard curve was prepared using different concentrations of MDA, and the extent of lipid peroxidation was calculated.
Nitric oxide (NO) production in the liver tissue
The NO contents of liver tissue were measured according to the method of Schmidt et al. [25]. The liver tissues were homogenized with a homogenizer by adding a physiological saline solution (0.9% NaCl) and centrifuged at 3,000 rpm for 10 min. After, 150 µL of the supernatant and 130 µL of distilled water were mixed, and then 100 µL of the mixture was added to the Griess reagent (1:1 ratio) and incubated at room temperature for 15 min. The absorbance was measured at 540 nm. The standard curve was prepared by different concentrations of NaNO2 and the inhibition of NO production was calculated.
Western blot analysis
The liver tissues were homogenized with lysis buffer containing a protease inhibitor cocktail. The homogenates were centrifuged at 12,000 rpm for 20 min at 4°C. After the supernatant was collected, the protein concentration was determined using the Bio-Rad protein assay kit (Bio-Rad, Irvine, CA, USA). An equal amount of proteins were resolved on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). After electrophoresis, proteins were transferred to polyvinylidene difluoride (PVDF) membrane (Millipore, Burlington, MA, USA). The membrane was incubated with 5% skim milk for 50 min at room temperature and then washed with PBS-T. The membrane was incubated with a primary antibody overnight at 4°C and then washed with PBS-T. The primary antibodies used were: β-actin (1:1,000; Cell Signaling, Beverly, MA, USA), NF-κB p65 (1:500, Abcam, Cambridge, UK), iNOS (1:500, Merk, Kenilworth, NJ, USA), COX-2 (1:1,000, Cell Signaling, Beverly, MA, USA), IL-1β (1:500; Bioss Antibodies, Boston, MA, USA), SOD (1:500; Santa Cruz, Dallas, TX, USA), catalase (1:500; Santa Cruz), and GSH-Px (1:500, Santa Cruz). The membranes were incubated with secondary antibodies for 1 h. The immuno-complexes were visualized by enhanced chemiluminescence solution (ELPIS Biotech, Daejeon, Korea) and bands were visualized with a chemiluminescent imaging system (Davinci Chemi, Seoul, Korea).
Statistical analysis
Data are presented as mean ± SD. Results were assessed using one-way analysis of variance (ANOVA) and Duncan's test for multiple comparisons. Statistical significance was considered as P < 0.05.
Go to :
RESULTS
Body weight, food intake, and food efficiency ratio
Changes in body weight, the amount of daily food intake, and food efficiency ratio are presented in Table 2. These data were not significantly different among all experimental groups.
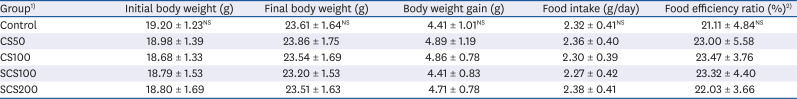
Table 2
Body weight and food intake in high cholesterol diet fed LDL receptor knockout mice for 10 weeks
Values are mean ± SD (n = 8 each group). Statistical analyses were conducted by Duncan's multiple range test (P < 0.05). NS, Non-significance.
1)Control, LDL receptor knockout (LDLR-KO) mice fed high cholesterol diet (HCD) and oral administration (OA) of purified water; CS50 and CS100, LDLR-KO mice fed HCD and OA of CS at a concentration of 50 and 100 mg/kg/day, respectively; SCS100 and SCS200, LDLR-KO mice fed HCD and OA of SCS at a concentration of 100 and 200 mg/kg/day, respectively.
2)Food efficiency ratio (%) : (total weight gain/total food intake) × 100.
![]()
Serum lipid profiles, atherogenic index, and cardiovascular risk index
As shown in Fig. 1, serum lipid profiles, atherogenic index (AI), and cardiovascular risk index (CRF) were determined. The administration of CS50, CS100, SCS100, and SCS200 significantly decreased the TG levels to 3.64 mmol/L, 3.72 mmol/L, 4.03 mmol/L, and 4.09 mmol/L, respectively, compared to those of the untreated control group (4.97 mmol/L). The concentration of TC was 71.45 mmol/L in the control group, whereas the CS50, CS100, SCS100, and SCS200 groups significantly reduced the TC level to 65.71 mmol/L, 64.20 mmol/L, 64.93 mmol/L, and 67.03 mmol/L, respectively. The LDL-C concentrations were significantly reduced in the CS50, CS100, SCS100, and SCS200 groups to 56.75 mmol/L, 54.67 mmol/L, 53.69 mmol/L, and 50.77 mmol/L, respectively, relative to that in the control group (63.96 mmol/L). The HDL-C concentrations were significantly increased in the CS50, CS100, SCS100, and SCS200 groups to 8.71 mmol/L, 9.74 mmol/L, 10.59 mmol/L, and 15.24 mmol/L, respectively, compared with the control group (4.07 mmol/L). Subsequently, the CS or SCS significantly decreased the AI and CRF. Compared with the control group, the CS50, CS100, SCS100 and SCS200 groups showed a decrease in the AI index by 6.98%, 6.22%, 5.35%, and 3.53%, and a decrease in the CRF index by 7.98%, 7.22%, 6.35%, and 4.53%, respectively.
 | Fig. 1Effects of chondroitin sulfate from shark cartilage (CS) and chondroitin sulfate from skate cartilage extract (SCS) on serum lipid profiles in high cholesterol diet fed LDL receptor knockout mice for 10 weeks. (A) TG, triglyceride; (B) TC, total cholesterol; (C) LDL-C, low density lipoprotein cholesterol; (D) HDL-C, high density lipoprotein; (E) AI, atherogenic index; (F) CRF, cardiac risk factor. See the legend of Table 1 for experimental groups in detail. Values are means ± SD (n = 8 per group). a~cMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
|
Serum inflammatory cytokine levels
The effects of CS and SCS on serum inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and PGE2 are shown in Fig. 2. The concentrations of TNF-α were decreased to 20.38 pg/mL, 18.08 pg/mL, 16.54 pg/mL, and 21.62 pg/mL in the CS50, CS100, SCS100, and SCS200 groups, respectively, compared with control group (26.46 pg/mL). In particular, the SCS100 group showed a significantly decreased level of TNF-α. In CS50, CS100, SCS100, and SCS200 groups, the IL-1β levels were significantly reduced to 124.53 pg/mL, 121.16 pg/mL, 129.47 pg/mL, and 126.87 pg/mL, respectively, from those in the control group (145.05 pg/mL). In addition, the IL-6 level in the CS50, CS100, SCS100, and SCS200 groups were decreased by 112.69 pg/mL, 119.19 pg/mL, 116.94 pg/mL, and 111.94 pg/mL, respectively, compared with control group (123.69 pg/mL). However, no significant difference in PGE2 levels was noted in any experimental group.
 | Fig. 2Inflammatory cytokine concentrations in high cholesterol diet fed LDL receptor knockout mice for 10 weeks. (A) TNF-α, tumor necrosis factor-alpha; (B) IL-β, interleukin-1beta; (C) IL-6, interleukin-6; (D) PGE2, prostaglandin E2. See the legend of Table 1 for experimental groups in detail. Values are means ± SD (n = 8 per group). a~cMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test. NS, Non-significance.
|
Inflammation-related protein expression in the liver
The effects of CS and SCS on the hepatic inflammatory-related protein levels, such those of as NF-κB, iNOS, COX-2, and IL-1β, are shown in Fig. 3. The administration of CS or SCS lowered the protein expression levels of NF-κB, iNOS, COX-2, and IL-1β. In particular, compared to those of the control group, the protein expression levels of NF-κB and IL-1β in the CS100, SCS100, and SCS200 groups were significantly decreased by 31.17%, 33.50%, and 42.03%, respectively, and by 13.35%, 19.88%, and 16.97%, respectively (P < 0.05). Those levels of iNOS in the CS50, CS100, SCS100, and SCS200 groups were significantly reduced by 24.72%, 51.69%, 53.61%, and 38.46%, respectively (P < 0.05). Those levels of COX-2 in the CS100 and SCS200 groups were significantly decreased by 49.00% and 41.44%, compared to those of the control group (P < 0.05)
 | Fig. 3Effects of chondroitin sulfate from shark cartilage (CS) and chondroitin sulfate from skate cartilage extract (SCS) on inflammation related protein expression in the liver of high cholesterol diet fed LDL receptor knockout mice. (A) Representative Western blot analysis; (B) NF-κB, nuclear factor kappa B; (C) iNOS, inducible nitric oxide synthase; (D) COX-2, cyclooxygenase-2; (E) IL-1β, interleukin-1beta. See the legend of Table 1 for experimental groups in detail. Values are means ± SD (n = 8 per group). β-actin was used as a loading control. a~eMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
|
Lipid peroxidation and nitric oxide formation in the liver
The inhibitory effects of CS and SCS on lipid peroxidation and NO are shown in Fig. 4. The hepatic MDA levels were significantly decreased in the CS50, CS100, SCS100, and SCS200 groups, to 8.04 µmol/mg protein, 5.38 µmol/mg protein, 6.33 µmol/mg protein, and 8.39 µmol/mg protein, respectively, compared with the control group (12.04 µmol/mg protein). In particular, the CS100 and SCS100 groups showed the lowest MDA levels. The NO levels were significantly reduced in the CS100, SCS100, and SCS200 groups by 124.81 µmol/mg protein, 123.09 µmol/mg protein, and 123.94 µmol/mg protein, respectively, but there was no significant difference, compared with that of the control group (130.55 µmol/mg protein).
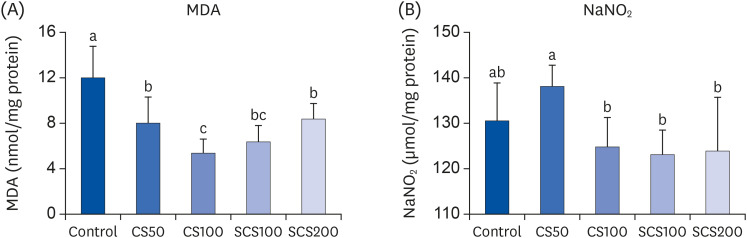 | Fig. 4Effects of chondroitin sulfate from shark cartilage (CS) and chondroitin sulfate from skate cartilage extract (SCS) on lipid peroxidation and nitric oxide formation in the liver of high cholesterol diet fed LDL receptor knockout mice. (A) MDA, malondealdehyde; (B) NaNO2. See the legend of Table 1 for experimental groups in detail. Values are means ± SD (n = 8 per group). a~cMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
|
The effects of CS and SCS on hepatic antioxidant enzymes, including SOD, catalase, and GSH-Px are shown in Fig. 5. The protein expression levels of SOD and catalase were significantly higher in the SCS100 (by 102.67% and 115.07%) and SCS200 groups (by 112.46% and 134.49%) than the control group. In addition, GSH-Px expression level was significantly increased in the CS50, CS100, SCS100, and SCS200 groups by 91.23%, 128.70%, 148.66%, and 174.12%, respectively, compared to that of the control group. In particular, SCS-administered groups showed a dose-dependent increase in the levels of hepatic antioxidant enzymes, including SOD, catalase, and GSH-Px.
 | Fig. 5Effects of chondroitin sulfate from shark cartilage (CS) and chondroitin sulfate from skate cartilage extract (SCS) on antioxidant enzymes expression in the liver of high cholesterol diet fed LDL receptor knockout mice. (A) Representative Western blot analysis; (B) SOD, superoxide dismutase; (C) Catalase; (D) GSH-Px, glutathione peroxidase. See the legend of Table 1 for experimental groups in detail. Values are means ± SD (n = 9). a~eMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
|
Go to :
DISCUSSION
Blood TG and TC are essential components for maintaining the function of the human body [26]. However, it has been reported that excessive intake of HCD increases the fat concentration in the blood, which causes atherosclerosis and NAFLD [156782728]. Therefore, it is crucial to control the blood lipid level in order to prevent these chronic diseases. LDLR-KO mice have widely used in the study for hyperlipidemia and hepatic lipid metabolism [42930]. Furthermore, the intake of HCD in LDLR-KO mice induces a rapid increase in plasma lipid levels as well as stimulates the TG synthesis in the liver. Chondroitin sulfate from salmon nasal cartilage has anti-hyperlipidemia via inhibition of absorption of dietary fat in high-fat diet fed mice [31]. In our previous study, SCS downregulated the protein expression levels of transcription factors, such as SREBP-1 and -2, thereby reducing serum TG and TC levels in LPS-injected mice [18]. In the present study, the CS and SCS administered groups had improved lipid profiles by decreasing the levels of TG, TC, and LDL-C, and increasing the levels of HDL-C compared with the control group. In particular, the AI and CRF indexes were the lowest in the SCS200 group among all experimental groups, which was probably due to the augmented HDL-C levels. These results might be associated with the reduced synthesis of fatty acid and cholesterol by SCS. These data indicate that SCS is expected to effectively alleviate hyperlipidemia in HCD-induced dyslipidemia.
Inflammatory responses are observed under the hyperlipidemia. Several inflammatory cytokines and mediators are involved in inflammation, such as TNF-α, IL-1β, IL-6, and PGE2 [32]. TNF-α is a major pleiotropic cytokine that exerts proinflammatory effects in atherosclerosis [32]. In addition, IL-6 and IL-1β are multifunctional cytokines that regulate various aspects of the immune response, acute-phase reaction, and hematopoiesis [33]. In addition, NF-κB plays a key role in hepatic inflammation. NF-κB activated by IκB phosphorylation leads to release numerous pro-inflammatory genes such as iNOS and COX-2 in the nucleus [3435]. iNOS leads to the production of excessive amounts of NO in the vascular smooth muscle cells, macrophages, and other cells [36]. COX-2 converts arachidonic acid to prostaglandin H2, which may trigger and maintain the inflammatory response in the various diseases, including atherosclerosis [37]. In the current study, the administration of SCS or CS decreased the serum levels of TNF-α and IL-1β and the expression levels of hepatic NF-κB, iNOS, and IL-1β, compared with those of the control group. In addition, the serum IL-6 level was reduced in the only SCS group. In particular, serum TNF-α and hepatic iNOS, COX-2, and IL-1β levels were the lowest in the SCS-administered group, with 100 mg/kg BW. In contrast, the serum IL-6 level was significantly lower only in the CS groups. These results were consistent with the previous study that chondroitin sulfate reduced serum inflammatory cytokines such as IL-1β and TNF-α in the obese mice and suppressed the activation of NF-κB in the inflamed human coronary artery endothelial cells [16]. In addition, chondroitin sulfate inhibits inflammatory reaction through downregulation of monocyte chemoattractant protein-1 (MCP-1) in the LPS-treated 3T3-L1 adipocytes [38]. Bovine-derived chondroitin sulfate attenuates IL-1β [39] or lipopolysaccharide [17] induced-inflammatory responses via suppression of NF-κB nuclear translocation. Similarly, our previous study demonstrated that SCS suppressed LPS-induced inflammation via downregulation of TNF-α, iNOS, and COX-2 in the liver tissue [18]. These results suggest that SCS could attenuate the inflammation induced by excessive intake of cholesterol in the serum and liver tissue, in a manner comparable to CS.
Oxidative stress elevated under hyperlipidemia is caused by free radicals produced by macrophages and directly damages lipid, protein, and other biological molecules. By contrast, oxidative stress could be accelerated due to hyperlipidemia and hypercholesterolemia [4041]. The MDA, the final product of lipid peroxides, affects cell membrane function, modifies protein, induces DNA damage, and reduces their function [42]. The excessive production of NO easily reacts with reactive oxygen species, which generates peroxynitrite in the human body [43]. The production of peroxynitrite leads to lipid peroxidation as well as remarkable modifications in the antioxidant defense system [44]. Oxidative stress accompanies a lower level of antioxidant enzymes [45]. Hypercholesterolemia diminishes the antioxidant defense system and inhibits the activities of antioxidant enzymes such as SOD, GSH, and catalase [44]. In the present study, CS or SCS-administered groups showed a significant reduction of hepatic MDA levels, compared to the control group. In contrast, the expression level of GSH-Px was significantly higher in all CS and SCS groups, when compared to the CS-administered group. Moreover, those levels of SOD and catalase were elevated in SCS groups in which effects were dose-dependent. These results suggested that the reduction of MDA level in the liver was partially attributed to the elevation of antioxidant enzymes. These results are consistent that SCS exhibited the protective effects against hepatic oxidative stress via up-regulation of GPx in LPS-injected mice [18]. Similarly, the previous studies reported the protective effects of chondroitin sulfate on oxidative stress by downregulation of extracellular ROS and upregulation of antioxidant-related proteins such as heme oxygenase-1 in the hydrogen peroxide-treated cells [17]. In addition, chondroitin sulfate attenuated oxidative stress through the reduction of lipid peroxidation and activation of antioxidant enzymes in the CCl4-induced oxidative stress mouse model [46]. Therefore, the results of the present study indicate that CS or SCS may contribute to alleviating the oxidative stress induced by HCD through the enhanced expression of antioxidant enzymes in hypercholesteremic mice.
In conclusion, we investigated the beneficial effects of SCS on hyperlipidemia-induced inflammation and oxidative stress and compared them with those of CS in HCD-fed LDLR-KO mice. Administration of SCS improved the levels of serum lipid, reduced the levels of serum and hepatic inflammation-related proteins, and augmented the antioxidant status in dyslipidemic mice. These effects of SCS were comparable with those of the CS group. Our findings suggest that the biological effects of SCS are attributed to the alleviation of hyperlipidemia, inflammation, and oxidative stress like shark CS.
Go to :
 XML Download
XML Download