

 PDF
PDF Citation
Citation Print
Print



The Sulwon Award for Scientific Achievement is the Korean Diabetes Association's highest scientific award and honors an individual who has excellently contributed to the progress in the field of diabetes and metabolism. The Sulwon Award is named after an emeritus professor, Eung Jin Kim, who founded Korean Diabetes Association. Prof. Minho Shong received the 11th Sulwon Award at 32nd Spring Congress of Korean Diabetes Association, May 9 to 11, 2019 at Gyeongju, Korea.
INTRODUCTION
Growth differentiation factor 15 (GDF15) was originally classified as a divergent member of the transforming growth factor beta (TGF-β) superfamily. GDF15 was initially discovered in 1997 through subtraction cloning, and named macrophage inhibitory cytokine-1 (MIC-1) due to increased secretion in response to proinflammatory cytokines, thus limiting the further activation of late phase macrophages [1]. A second group identified a gene in response to nonsteroidal anti-inflammatory drugs (NSAID), and defined GDF15 as NSAID-activated gene (NAG-1) [2]. GDF15 is also known as placental transformation growth factor-β (PTGF-β) and prostate derived factor (PDF) [34]. Although initially classified as a member of the TGF-β superfamily, the recent discovery of the glial cell line-derived neurotrophic factor (GDNF) family receptor α-like (GFRAL) protein as the cognate receptor of GDF15 has led to attempts to reclassify GDF15 as a member of the GDNF family [5].
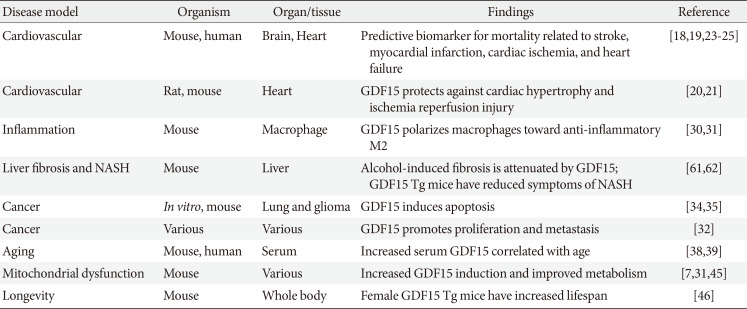
GDF15 is highly expressed in the placenta and during pregnancy. Whole body knockout of GDF15 in mice showed normal development and fertility, but mice were more prone to diet-induced obesity due to spontaneous food intake, suggesting that GDF15 atypically regulates appetite [6]. Cellular stress is known to highly upregulate GDF15 in both humans and mice [7891011]. Plasma concentrations of GDF15 increase during metabolic adaptations such as exercise, and in various pathologies including cardiovascular disease, inflammation, cancer, obesity, mitochondrial disease, as well as during aging [11121314151617]. GDF15 induction in various diseases is summarized in Table 1.
INDUCTION OF GDF15 UNDER VARIOUS CONTEXTS
Cardiovascular disease
Many of the early studies on GDF15 have focused on its role in cardiovascular disease. GDF15 is a predictive biomarker for cardiovascular risk as well as all-cause mortality [18192021]. Serum GDF15 increases in various conditions such as ischemia, myocardial infarction, heart failure, and stroke. In particular, myocardial infarction was found to elevate the expression of GDF15 in response to increased reactive oxidative species and pro-inflammatory cytokines [22232425]. Common to these studies was that GDF15 was a strong predictor of mortality. However, it should be noted that increased GDF15 as a predictive biomarker does not establish causation. Additionally, there are conflicting reports as to whether GDF15 protects against or exacerbates cardiovascular disease [2023]. Thus, these studies should be approached with discretion. GDF15 in relation to cardiovascular disease has been previously reviewed by Wang et al. [26].
Inflammation
GDF15 has also been reported to have an anti-inflammatory effect [2728]. Immune balance is maintained by pro-inflammatory type 1 and anti-inflammatory type 2 immune responses [29]. Our group found that GDF15 induces an anti-inflammatory type 2 response through M2 polarization of macrophages [30]. In our study, we generated a macrophage-specific mouse model with genetic ablation of the CR6-interacting factor 1 (Crif1) gene, which is essential for the biogenesis and assembly of the mitochondrial oxidative phosphorylation complex. Macrophage-specific Crif1 deletion mice showed reduced GDF15 induction and increased polarization to pro-inflammatory M1 macrophages. M2 polarization by GDF15 was also observed in a separate study on adipocyte-specific Crif1 knockout mice [31]. Increased expression of GDF15 was observed in adipose tissue of Crif1- deleted mice, which was accompanied by an increase in M2 polarization of macrophages. Double knockout of Crif1 and GDF15 abrogated M2 polarization and increased the proportion of M1 macrophages. In addition, Luan et al. [15] observed that the expression of GDF15 in humans and mice increased during inflammatory conditions caused by bacterial and viral infections, and revealed that GDF15 induces cardioprotective effects during acute inflammation. Collectively, these results indicate that GDF15 can have an anti-inflammatory effect.
Cancer
Significant increases of GDF15 expression have been observed in various types of cancers including colon, breast, pancreas, liver, thyroid, and ovarian, and can have both a pro- and anti-tumorigenic effect [1232]. The expression of GDF15 is regulated by p53, early growth response protein 1 (EGR-1), mitogen activated protein kinase (MAPK), or the AKT/glycogen synthase kinase 3 beta (GSK-3 beta) pathways in tumor microenvironments [33]. GDF15 is heavily involved in cancer progression through proliferation, invasion/metastasis, and drug resistance. In most cancer types, GDF15 induction promoted tumor proliferation as well as invasion/metastasis via the p38 MAPK, AKT, and ERK-1/2, SMAD 2/3, and GSK-3 beta pathways [32]. In contrast to its pro-tumorigenic role, GDF15 was also shown to inhibit cancer by inducing apoptosis in lung cancer and glioma [3435]. In these studies, the apoptotic role of GDF15 was revealed by using GDF15 transgenic mice and cell lines overexpressing GDF15. Cell apoptosis in lung cancer and glioma was activated by the p38 MAPK and PI3K/AKT signaling pathways, respectively. Therefore, it is likely that the dichotomous role of GDF15 in cancer can be explained by its anti-tumorigenic response to limit tumor growth in the initial stages of tumorigenesis. However, in the later stages, cancer cells may exploit GDF15 as a means to escape immune surveillance, and thus promote a more pro-tumorigenic environment conducive to proliferation and metastasis.
Aging
Aging is characterized by a gradual decline in cellular homeostasis [36]. As aging progresses, not only do the bodily functions weaken, but inflammation and tissue injury become more common. As GDF15 is known to be induced by stress, it is unsurprising that plasma GDF15 increases with age [163738]. A recent study utilized aptamer-based proteomics to conduct a large-scale study of plasma proteins in human cohorts [39]. A total of 2,925 plasma proteins from 4,263 subjects between the ages of 18 and 95 years were analyzed. GDF15 was found to be a significant marker for aging and showed undulating increases based on age. Furthermore, GDF15 was found to be highly conserved between mice and humans, suggesting that GDF15 is an ideal therapeutic target for aging research. A more recent study attempted to find proteomic signatures that changed through a 9-year follow-up from older adults [40]. As mobility disability is an indicator of health in the elderly, a method of comparing the association between mobility and plasma proteins was implemented. In this study, the authors found that GDF15, plasma cathepsin S (CTSS), and thrombospondin-2 (THBS2) are novel biomarkers associated with mobility disability in the elderly.
Mitochondrial dysfunction
Mitochondrial dysfunction has been implicated in various diseases ranging from metabolic disorders to cancer and aging-related disorders, and is a common underlying factor which may account for GDF15 induction in various pathophysiologic contexts [4142]. GDF15 is considered as a mitochondrial cytokine (mitokine), and induced by mitochondrial dysfunction [783738]. The concept of mitokine was first mentioned in the C. elegans model, where upregulation of the mitochondrial unfolded protein response (UPRmt) in the worm nervous system resulted in the distal activation of the UPRmt in the gut [43]. In the context of mammalian systems, the increased complexity of the organism results in a differing response where mitokines can regulate systemic metabolism [7104445]. In lower organisms, the UPRmt is reported as an important factor in longevity [43]. By contrast, in mammalian systems, the UPRmt confers a metabolic benefit in the form of improved glucose tolerance and insulin sensitivity.
Fibroblast growth factor 21 (FGF21) and GDF15 are considered as mammalian mitokines, which can have a metabolic benefit. Further investigations will be required to confirm whether elevated mitokine induction can lead to increased lifespan in higher organisms. Several murine studies and correlative analyses in human patients suggest that mitokines can in fact increase the longevity in mammals [374647]. Transgenic mice overexpressing FGF21 were reported to have a 36% increase in median lifespan. These benefits were attributed to decreased growth hormone/insulin-like growth factor-1 (IGF-1) signaling in the liver [47]. Another study in transgenic mice overexpressing hNAG-1/GDF15 found that female mice had increased mean and median lifespan, with improved metabolic parameters including lower body weight, improved glucose tolerance and insulin sensitivity, and decreased serum levels of insulin and IGF-1. The lifespan-enhancing effect was more pronounced when mice were fed a high fat diet.
While FGF21 initially received significant interest, a human clinical trial of the FGF21 analog, LY2405319, failed to recapitulate the glucose-lowering effects observed in rodents and non-human primates [48]. It remained to be seen whether GDF15 could also be a pharmacological agent which can be used in the treatment of metabolic diseases. However, the recent discovery of the cognate GDF15 receptor has spurred a new interest in GDF15 as a novel therapeutic drug.
GFRAL AND THE METABOLIC ROLE OF GDF15
Historically, the studies showing high expression and serum levels of GDF15 in various cancers attempted to identify the receptor and downstream signaling mediating the biological action of GDF15 [20495051525354]. As GDF15 is a distant member of the TGF-β superfamily, most in vitro studies initially suggested that treatment of recombinant GDF15 (rGDF15) can phosphorylate SMAD2/3 dose-dependently through the TGF-β receptor type I/II [495152] or avian erythroblastosis oncogene B (ErbB2/HER2) [53]. However, recent studies have revealed that purified rGDF15 prepared from mammalian cells also included a small amount of TGF-β1, which caused the phosphorylation of SMAD2 [55].
GFRAL, an orphan member of the GDNF receptor family, was newly identified in 2017 as the receptor for GDF15 by four independent research groups from the pharmaceutical industry [5565758]. Through the screening of a cDNA library including approximately 4,000 cell-surface proteins, the studies found that GFRAL strongly interacted with GDF15; the mutation of valine 87 and isoleucine 89 resulted in a loss of the binding affinity of GDF15 with GFRAL, while tryptophan 32 was found to be critical for the inhibition of food intake [57]. RET, a tyrosine kinase coreceptor, was necessary for the GDF15-GFRAL signal transduction and downstream activation of AKT, ERK1/2, and phospholipase C-γ (PLCγ) in neuroblastoma cell lines transfected with GFRAL [5758]. A study using cryogenic electron microscopy (cryo-EM) confirmed the binding interaction in the GDF15-GFRAL-RET complex, in which interaction is formed between the helix-loop-helix region of GFRAL and the cadherin-like domain (CLD) 1/2 of RET. GDF15 binds to the cysteine-rich domain (CRD) of RET [59].
The expression of GFRAL in rodents, non-human primates, and humans is highly restricted to the area postrema (AP) and nucleus of the solitary tract (NTS) of the hindbrain. The representative physiological action of the GDF15-GFRAL axis resulted in an anorexigenic effect, causing a reduction in body weight [54956575860]. Prior to the identification of GFRAL, several studies already reported that administration of GDF15 triggered the activation of c-Fos in the AP and NTS regions, resulting in reduced food intake and decreased body weight in a leptin-independent manner [4960]. Consistent with previous results, food intake and body weight were reduced in control mice administrated a single or chronic dose of rGDF15 (0.1–0.3 mg/kg), but these effects were completely neutralized in Gfral knockout mice [5565758]. More recently, it was reported that administration of rGDF15 acutely (0.01 to 0.1 mg/kg) caused conditioned taste aversion (CTA) similar to taste aversion with lithium chloride, which was consistent with observations showing that GDF15 also activated c-Fos in the parabrachial nucleus (PBN) to relay the aversive signals [511]. Thus, recent studies have established the main role of GDF15 as orchestrating food intake and body weight.
Most TGF-β family members are dimeric polypeptides and serum GDF15 was detected in its mature form, which is consistent with the observation of the binding structure via crystallography and cryo-EM [1013]. Despite the clear interaction between GDF15 and GFRAL, there are still several unanswered questions pertaining to possible peripheral activity.
PERIPHERAL ACTIVITY OF GDF15
Several lines of evidence support direct GDF15 activity in the periphery which cannot be accounted for by reduced appetite. As GFRAL expression is limited to the AP and NTS region of the hindbrain, it is highly possible that GDF15 may act directly through an as-of-yet undiscovered mode of action. One study observed that in a pediatric model of heart disease via knockout of cardiomyocyte-specific estrogen-related receptor alpha (ERRα) and gamma (ERRγ) in mice, serum GDF15 was increased by 4-fold by day 10. Growth hormone levels in the serum were unchanged; however plasma IGF-1 levels were reduced, suggesting impaired liver growth hormone signaling. This resulted in the diminished body growth of mice [17].
We and others have found that increased serum GDF15 from the liver can reduce the symptoms of nonalcoholic steatohepatitis (NASH) and liver fibrosis [6162]. Liver-specific transgenic mice expressing human GDF15 were resistant to methionine-choline deficient (MCD) diet-induced NASH. No difference in body weight or feeding behavior was observed between the control group and transgenic mice on the MCD diet, suggesting that human GDF15 can have a direct anti-inflammatory effect on the liver immune environment [62]. In the liver fibrosis model, CCl4 and alcohol treatment in mice increased GDF15 induction in hepatocytes, which ameliorated liver fibrosis, as GDF15 knockout mice had exacerbated liver fibrogenesis. Furthermore, GDF15 knockout mice had increased TNF-α production, activated CD4+ and CD8+ T cells, and infiltration of monocytes and neutrophils [61]. Taken together, these two studies demonstrate that GDF15 can have a direct effect on the liver immune environment which cannot be explained through GFRAL-mediated feeding behavior.
One possible explanation for peripheral GDF15 activity can be gleaned from cancer studies. Early studies have found that various forms of GDF15 were secreted, which included not only the dimeric mature GDF15 (aa 197–308, approximately 30 kDa), but also the monomeric pro-GDF15 (aa 29–308, approximately 40 kDa), dimeric pro-GDF15 (approximately 80 kDa), and even the pro-peptide GDF15 (aa 29–196, approximately 28 kDa) [16364]. Although mature GDF15 was detected in the serum of a xenograft cancer model [63], pro-GDF15 was more rapidly secreted than mature GDF15 in transfected monocytoid cells and locally stored to mediate the biological activity of GDF15 [65]. Many of the previous studies focused on mature GDF15 in the context of pathological conditions such as cancer, cachexia, inflammatory disease, and type 2 diabetes mellitus. Thus it remains to be seen whether the various forms of GDF15 can exist in the serum and have any peripheral activity. This viewpoint is discussed in more detail by Baek and Eling [12].
Despite some evidence of peripheral GDF15 activity, in the absence of the identity of its cognate peripheral receptor, currently it is difficult to ascertain whether GDF15 can have a direct peripheral effect, thus necessitating further studies in this area. This problem is however exacerbated by commercially available rGDF15 contamination with TGF-β1 and endotoxin levels which may invalidate previous in vitro studies. The generation of rGDF15 in yeast-based systems may facilitate further studies on GDF15 signaling pathways in peripheral organs.
THERAPEUTIC APPLICATIONS
GDF15 can both reduce food intake and increase energy expenditure [4958]. GDF15 acts through a brain-stem-restricted receptor, through which it reduces food intake and lowers body weight. Metformin, the world's most prescribed anti-diabetic drug, is effective in preventing type 2 diabetes mellitus in people with high risk. Metformin elevates circulating levels of GDF15, which is necessary to obtain its beneficial effects on energy balance and body weight [66]. Metformin likely elicits a GDF15-independent effect on circulating levels of glucose and insulin by primarily increasing weight loss, secondary to anorexia. GDF15 levels do increase following sustained high-fat feeding or dietary amino acid imbalance in mice [11]. Mice fed a lysine-deficient diet activate the integrated stress response, which increases serum GDF15 levels. Pharmacological GDF15 administration to mice triggers CTA in response to nutritional stress, establishing the GDF15-GFRAL axis as critical to stress pathway-induced weight loss.
CONCLUSIONS
Gdf15 is a stress-related protein which is elevated in various pathologies. We have consistently found that Gdf15 is induced in mitochondrial defect models which generally show increased energy expenditure and resistance to high fat diet-mediated weight gain and glucose intolerance. With the discovery of GFRAL, it is indisputable that GDF15 regulates feeding behavior. However, several studies consistently show that GDF15 can have a peripheral activity. The lack of a definite peripheral Gdf15 receptor necessitates the need to identify a peripheral receptor or alternate mode of action which can mediate these peripheral responses. Increased availability of rGDF15 through improved production methods will likely facilitate further research in this area.
 XML Download
XML Download