

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Osteoporosis is a skeletal disorder characterized by compromised bone strength [1]. Fractures resulting from osteoporosis are a worldwide public health problem, causing substantial disability, morbidity, and mortality in the elderly [23]. Because of the global increase in the size of aging populations, osteoporotic fracture (OF) is expected to further impose considerable economic burden on health care systems [4]. Although bone strength is a concept that integrates bone mass and bone quality [5], the current operational definition of osteoporosis only relies on bone mineral density (BMD) due to the lack of clinically feasible methods of measuring bone quality [6]. Low BMD assessed by dual-energy X-ray absorptiometry is an important risk factor for OF development [7], but there is increasing concern about the ability of BMD to predict OF. Epidemiological cohort studies demonstrated that up to half of incident fractures occurred in individuals with BMD T score above −2.5, the diagnostic cut-off defining osteoporosis. To overcome the low sensitivity of BMD measurements, a World Health Organization working group developed a fracture risk assessment tool (FRAX) based on relevant clinical risk factors (CRFs) with or without BMD [8]. The CRFs, selected from the results of large prospective studies, include sex, age, weight, height, previous and parental fracture history, lifestyle factors, glucocorticoids, and the presence of secondary osteoporosis [8]. Although FRAX has markedly contributed to the improved prediction of fracture risk, the overall ability of FRAX to identify individuals at high risk for OF remains suboptimal [9]. Efforts have therefore been made to identify potential biomarkers that can predict the risk of OF, independent of or combined with BMD and CRFs.
Go to : 
FRACTION PREDICTION USING CONVENTIONAL BONE TURNOVER MARKERS
Bone is a metabolically active tissue that undergoes continuous turnover through sequential phases of osteoclastic bone resorption and osteoblastic bone formation [10]. The ultimate aims of this finely controlled process are the preservation of mineral homeostasis and mechanical characteristics. Bone turnover markers (BTMs) reflecting the rate of bone formation or the rate of bone resorption may be clinically useful for osteoporosis management [1112]. Based on a comprehensive analysis of blood and urinary markers related to bone remodeling, the International Osteoporosis Foundation and the International Federation of Clinical Chemistry have designated serum procollagen type I N-propeptide and serum C-terminal telopeptide of type I collagen (CTX) as reference standards for bone formation and bone resorption, respectively [131415]. Although BTMs are utilized clinically to monitor the short-term efficacy of anti-osteoporotic treatment [16], several prospective studies have suggested that BTMs may be predictors of future fracture risk [17]. For example, the Epidémiologie de l′Ostéoporose (EDIPOS) [18] and Os des FEmmes de LYon (OFELY) [19] studies of postmenopausal women reported that high levels of bone formation markers (BFMs) and/or bone resorption markers (BRMs) were significantly associated with high OF risk, independent of BMD. A recent systematic review and meta-analysis also highlighted the potential role of BTMs in predicting future fractures [20]. In most clinical studies, however, the relative risk of fracture of these BTMs increased approximately 2-fold per standard deviation (SD) or is in the highest quartile [18192122], making the ability of BTMs to predict future fracture risk insufficient. In addition, other longitudinal studies do not support the clinical utility of BTMs in assessing fracture risk in older women and men [2324]. Collectively, these studies showed that the contribution of BTMs to the prediction of OF risk remains inconclusive.
Go to :
EMERGING BIOMARKERS OF BONE METABOLISM
Increased understanding of the regulatory mechanisms underlying skeleton homeostasis has led to the identification of novel biological markers of bone metabolism. This review highlights potential biomarkers predictive of human bone health and their clinical applicability.
Sphongosine-1-phosphate
Sphongosine-1-phosphate (S1P), a highly bioactive lysophospholipid, which can function as both an extracellular mediator and an intracellular second messenger, has been shown to be involved in diverse biologic processes, including the migration, localization, apoptosis, proliferation, and differentiation of various cell types [25]. This pleiotropic molecule has also been found to play multiple roles in bone remodeling. Both osteoblasts and osteoclasts express S1P receptors (S1PRs), which belong to the G protein-coupled receptor family [2627]. S1P secreted by osteoclasts stimulates the motility, proliferation, and survival of osteoblasts [262829]. In addition, mice lacking osteoclast-specific cathepsin K exhibited higher bone formation in vivo through the enhancement of S1P production by osteoclasts [30]. Collectively, these findings indicate that S1P can act as an osteoclast-derived coupling factor, resulting in bone formation [10].
In contrast to its role in bone protection, S1P has different effects on osteoclastic biology. S1P in cocultures of osteoblasts and bone marrow (BM)-derived macrophages markedly potentiated osteoclastogenesis by stimulating receptor activator of nuclear factor-κB ligand (RANKL) secretion from osteoblasts [31]. Elevated circulating S1P levels in vivo induced the migration of osteoclast precursors from blood to bone with low S1P levels by a mechanism involving S1PR2-mediated chemorepulsion (i.e., negative chemotaxis away from S1P), and consequently facilitated bone resorption [2732]. Consistently, inhibition of S1PR2 in mice attenuated bone loss by reducing the number of osteoclasts attached to the bone surface [32]. The complex roles of S1P on skeleton, with both anabolic and catabolic activities, indicate a need to clarify the primary effects of S1P during bone remodeling.
To investigate whether the dominant effect of S1P in human bone metabolism consists of bone formation, resorption, or coupling, we firstly performed clinical study in 357 postmenopausal women with measurement of S1P and BTM [33]. After adjustment for potential confounders, plasma S1P concentrations were found to be positively associated with the levels of BRMs, such as urinary N-terminal telopeptide of type I collagen and serum CTX, but not with the levels of BFMs, such as serum bone-specific alkaline phosphatase and osteocalcin [33]. These results suggested that S1P primarily stimulates bone resorption rather than influences bone formation or coupling in humans. Furthermore, in a unique study using simultaneously obtained blood and BM samples, a higher blood/BM S1P ratio was found to be associated significantly with a greater risk of osteoporotic hip fracture [34]. Together, all these findings provide clinical evidence that the adverse effects of S1P on human bone homeostasis were attributable to the considerable S1P gradient between peripheral blood and BM, and to the resultant bone resorption via the chemorepulsion-induced increase of pre-osteoclasts in bone.
The interesting point regarding S1P is that this molecule is abundant in the circulation but low in other tissues due to its irreversible degradation by S1P phosphatase and/or S1P lyase [3536]. This easily measurable property in blood made us to hypothesize that S1P could be a candidate biomarker predictive of osteoporosis-related phenotypes. To investigate this possibility, we performed an age- and BMI-matched case-control study [37]. Plasma S1P concentrations were inversely correlated with BMD at various sites, as well as being 36.3% higher in patients with than in those without vertebral fractures (VFs) [37]. Higher circulating S1P levels were associated with higher risk and number of osteoporotic VFs, with statistical significance persisting even after adjustment for BMD [37]. A subsequent 3.5-year follow-up study demonstrated that high baseline blood S1P levels could be a potential predictor of high incident fractures [38]. Importantly, the association between blood S1P and fracture was perfectly replicated in another cohort. A recent prospective study with a 5.2-year follow-up period found that the hazard ratio for OF was 9.89-fold higher in women in the highest than in those in the lowest S1P quartile [39]. Taken together, these clinical studies consistently indicated that S1P may be a promising blood biomarker predictive of poor bone health outcomes.
Leucine-rich repeat-containing 17
Leucine-rich repeat-containing 17 (LRRc17) is a 37 kDa protein with secretary feature containing five putative LRR domains [40]. Previous experimental study well characterized the role of LRRc17 in bone metabolism as an inhibitor of RANKL-induced osteoclast differentiation [41]. Briefly, LRRc17, highly expressed in osteoblasts, decreased RANKL-mediated nuclear factor of activated T-cells, cytoplasmic 1 (NFATc1) signaling, followed by attenuating osteoclastogenesis from BM precursors [41]. These findings suggest that low circulating LRRc17 level may be a risk factor for OF. In fact, Hong et al. [42] demonstrated that postmenopausal women in the lowest plasma LRRc17 tertile had a 3.32-fold higher odds ratio (OR) for OF than those in the highest tertile and that each log-unit decrease in plasma LRRc17 concentration was associated with a 46% higher risk of OF after adjustment for bone mass and CRFs. Interestingly, the area under the receiver operating characteristics curve, integrated discrimination improvement, and category-free net reclassification improvement analyses consistently revealed that the addition of blood LRRc17 to a FRAX model markedly improved its ability to predict OF [42]. These findings suggest that LRRc17 is an independent and additive biomarker for OF in postmenopausal women.
Macrophage migration inhibitory factor
Macrophage migration inhibitory factor (MIF) is a 37.5 kDa cytokine exerting various biologic effects through its receptor CD74. In addition to its involvement in the regulation of inflammatory and immune processes [43], MIF has emerged as a key player in bone metabolism. For example, histomorphometric analyses showed that mice overexpressing MIF had osteoporotic phenotypes resulting from high bone turnover [44], whereas MIF-deficient mice were resistant to the increases in bone resorption and bone loss after ovariectomy [45]. These detrimental effects of MIF on bone homeostasis in humans were validated by investigating the association of blood MIF level with osteoporosis-related phenotypes in postmenopausal women [46]. After controlling for potential confounders, plasma MIF concentrations were found to correlate inversely with BMD at the lumbar spine and proximal femur, and to correlate positively with levels of BFM and BRM [46]. Each SD increase of blood MIF level was significantly associated with a multivariable-adjusted OR of 1.32 for the risk of osteoporosis [46]. These results suggest that circulating MIF may be a potential biomarker associated with human bone metabolism.
Sclerostin
The Wingless (Wnt) ligands are pivotally involved in bone metabolism through their interaction with a receptor complex consisting of seven-transmembrane frizzled-related proteins and low-density lipoprotein receptor-related protein 5/6 (LRP5/6) [47]. Activation of the Wnt signaling pathway induces the translocation of stabilized cytoplasmic β-catenin into the nucleus, subsequently stimulating the transcription of key genes involved in osteoblast differentiation [48]. Sclerostin competitively binds to LRP5/6 and thus acts as an inhibitor of the Wnt pathway, resulting in reduced osteoblastogenesis [49]. Sclerostin also stimulates the production of osteocyte-derived RANKL [50], indicating that sclerostin is a dual action molecule, having deleterious effects on skeletal homeostasis by increasing bone resorption as well as by reducing bone formation.
Sclerostin is a secreted protein, primarily by osteocytes, with its concentration increasing with age [51]. Importantly, circulating and BM plasma levels of sclerostin were strongly correlated, suggesting that blood sclerostin concentration may represent its local production in the bone microenvironment [52]. In view of these features, sclerostin has been expected as an ideal biomarker of human bone metabolism, and several epidemiologic studies have been performed to clarify this point. Consistent with its role as a negative regulator of bone remodeling, higher sclerostin concentrations in blood were a strong and independent risk factor for higher fracture occurrence in older Caucasian and Arab women [5354]. By contrast, the OFELY study of 572 postmenopausal women found no association between baseline serum sclerostin levels and incident fracture [55]. Moreover, other clinical studies showed that older men and women with high sclerostin levels were at reduced risk of fracture [5657]. Because of these discrepancies, sclerostin is not regarded as a predictor of fracture, and its measurement is not yet recommended in clinical practice.
RANKL
RANKL plays essential roles at every stage of osteoclastogenesis and bone resorption [58]. Although RANKL is expressed in various cell types, including osteoblasts, conditional genetic deletion indicates that osteocytes are the major source of RANKL needed for osteoclast formation [5960]. The development of sensitive assays to effectively measure RANKL in body fluids [61] led to clinical studies evaluating the association between circulating RANKL levels and metabolic bone diseases. These studies, however, yielded conflicting results [626364], and critical concerns were raised regarding the origin and nature of circulating RANKL. For example, serum RANKL concentrations were inversely correlated with RANKL mRNA expression in bone tissue [65]. Furthermore, Drake et al. [52] reported that RANKL could not be detected in most peripheral blood samples. Therefore, the ability of blood RANKL concentration to predict fracture remains unclear.
Periostin
Periostin, a glutamate-containing extracellular matrix protein, was originally isolated from murine pre-osteoblast MC3T3-E1 cells and initially designated osteoblast-specific factor 2 [66]. Subsequent research demonstrated that this protein was mainly expressed in the periosteum and was therefore renamed periostin. Results from genetically altered mice showed that periostin plays a beneficial role in bone metabolism [67]. For example, bone mass was lower and microarchitecture was weaker in periostin-deficient mice than in control mice, resulting in reduced bone strength in periostin-deficient mice [68]. The notable findings, demonstrated by Bonnet et al. [6869], were that mechanical loading markedly increased the expression of periostin in stimulated bone, and that these changes correlated with the improved biomechanical properties of long bones through the increase of bone-forming activity, especially at the periosteal site. These results suggest that periostin may be a distinctive biomarker of periosteal metabolism, a process that cannot be determined by conventional BTMs, most of which are biomarkers of endosteal bone remodeling [70].
Despite the clear biological importance of periostin in rodent skeleton, the relationship between blood periostin level and osteoporosis-related phenotypes in humans was determined only recently. In a 7-year prospective cohort of 607 postmenopausal women, the risk of incident fracture was 1.88-fold higher in the quartile with the highest than in that with the lowest circulating periostin concentration, independent of BMD and CRFs [71]. These results were replicated in another study, which showed that the OR per each increase in SD of blood periostin for any type of OF was 1.58 in the multivariable adjustment model [72]. Because periostin showed bone anabolic effects in experimental studies, these clinical data seemed counterintuitive. Although the reasons for these unexpected findings remain uncertain, Rousseau et al. [71] hypothesized that periostin expression may be increased to compensate for increased bone fragility. Although periostin expression after bone growth is maintained at a low level under normal conditions, this protein is re-expressed in response to pathologic mechanical strains in adults [67]. For example, periostin expression was markedly increased during the early stages of fracture healing, possibly contributing to periosteal callus formation [73]. Therefore, circulating periostin concentrations may be increased in subjects with poor bone strength and resultant fractures through a process involving increased mechanical stress on the remaining bones to maintain skeleton integrity.
Because periosteum is an outer membrane of cortical bone, the role of circulating periostin in terms of OF risk might be more prominent in long bones than in spine, which is mainly composed of trabecular bone. To answer this question, we analyzed non-VFs and VFs separately in a recent case-control study [72]. Interestingly, higher plasma periostin levels were significantly associated with a higher risk of non-VFs and lower femur BMD, but not with a higher risk of VFs and lower spinal BMD [72]. These results suggest that the effects of periostin on human skeleton may depend on bone types, and that this protein could serve as an effective biomarker, especially for non-VFs.
Other bone regulatory factors
In addition to the factors described above, dipeptidyl-peptidase 4 [74], slit guidance ligand 3 [1075], cathepsin K [76], dickkopf-related protein 1 (DKK1) [11], and fibroblast growth factor 23 [77] may act as circulating candidate markers of bone metabolism. Since most researchers in bone field have a consensus on the limitations of BMD in fracture risk assessment, novel biomarkers would continue to be identified, according to the increase of knowledge about bone regulation, and be validated for the clinical utility.
Go to :
BIOMARKER RISK SCORE ENHANCING THE ACCURACY OF FRACTURE PREDICTION
Because osteoporosis is a multifactorial disease involving various kinds of players, the contribution of any single biomarker to fracture prediction improvement may not be strong enough. This led to a formulation of a “biomarker risk score” by combining the available candidates using the optimal linear combination method [78] in a case-control control including 160 postmenopausal women as follow: −0.061×[(DKK1–mean)/SD]×β (coefficient)+0.212×[(MIF–mean)/SD]×β+0.378×[(LRRc17–mean)/SD]×β−0.635×[(periostin–mean)/SD]×β+0.636×[(sclerostin–mean)/SD]×β. The addition of a biomarker risk score to a model including CRFs and hip BMD was found to improve the ability to detect OF by 31.8% (area under the curve 0.62 vs. 0.82, P<0.001) (Fig. 1). This scoring system was cross-sectionally performed in relatively small samples, and thus could be applied to other populations. However, this preliminary analysis might provide important implications in that it suggests the necessity of integrating the established biomarkers for effective fracture risk assessment.
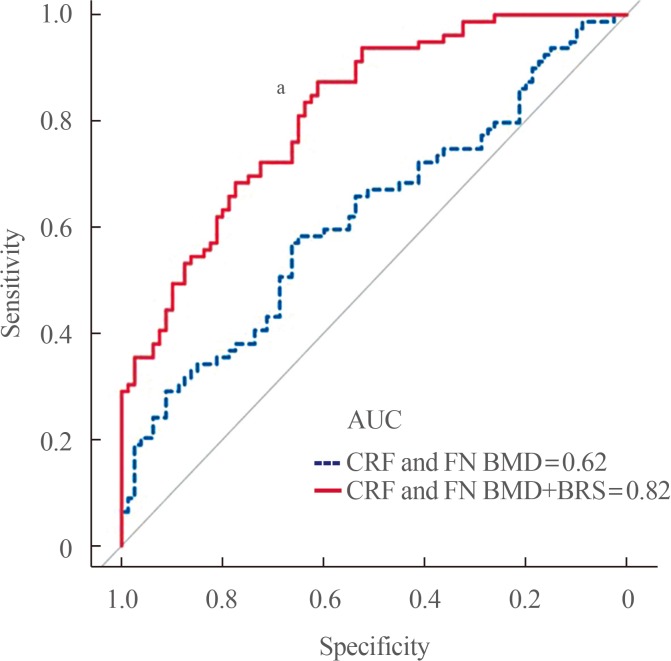 | Fig. 1Comparison of the receiver operating characteristics curves and the area under the curve (AUC) of clinical risk factor (CRF) and femur neck bone mineral density (FN BMD) with or without biomarker risk score (BRS) for detecting osteoporotic fracture in 160 postmenopausal women. aSignificant improvement of the AUC upon addition of BRS to CRF and FN BMD.
|
Go to :
POSSIBLE ROLE IN PREDICTING RESPONSES TO NEW ANTI-OSTEOPOROTIC MEDICATIONS?
Two new therapeutic agents have been introduced for the treatment of osteoporosis [7980]. One of these agents, romosozumab, is a dual acting drug that simultaneously stimulates bone formation and decreases bone resorption by inhibiting sclerostin [81]. The second agent, denosumab, is a fully human recombinant monoclonal antibody to RANKL, blocking the binding of RANKL to the RANK receptor on cells of the osteoclast lineage, thereby suppressing bone resorption [82]. Pivotal phase 3 clinical trials of both drugs demonstrated their efficacy in reducing the incidence of fracture [8384]. In particular, sequential treatment with romosozumab followed by denosumab resulted in a greater increase in bone mass than any other treatment strategies [85]. With the increasing importance of romosozumab and denosumab in the osteoporosis management, physicians may be interested in biomarkers that can predict their therapeutic response in advance. For example, it can be speculated that bone anabolic and/or anti-resorptive activities might be much stronger when romosozumab and denosumab are injected into patients with higher circulating pretreatment levels of sclerostin and RANKL, respectively. Unfortunately, as discussed earlier in this paper, clinical studies to date have not shown consistent results regarding the association between circulating sclerostin and bone parameters [5354555657], and the measurement of RANKL in blood was challenging due to its low concentration [52]. At present, therefore, existing evidence does not support the role of circulating sclerostin and RANKL as predictors of treatment responses to new anti-osteoporotic medications.
Go to :
CONCLUSIONS
Main objective of bone research is to reduce the burden of fragility fracture causing high disability and morbidity. To achieve this goal, the subjects vulnerable to fracture should be early detected. Assessment of fracture risk has been moving toward an absolute-risk approach that can be estimated from individual risk profiles, largely based on CRFs and BMD. Although the current model is used worldwide, there is a need improve the accuracy of fracture prediction. The present review described emerging biomarkers of bone metabolism and the importance of biomarker risk score, generated by combining individual biomarkers. Although many challenges, such as bone specificity, intra-individual and biologic variability, and cost-effectiveness, remain prior to applying these biomarkers to actual clinical practice, researchers in bone field are recognizing the limitations and endeavoring to minimize them. Therefore, future efforts, including the validation and replication in the longitudinal cohorts of various populations, are expected to increase clinical applicability of potential biomarkers for the fracture risk assessment.
Go to :
 XML Download
XML Download