

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Numerous recent studies have revealed the functions of high-density lipoprotein (HDL), which include roles in lipid transport, inflammation, and immunity.1) To date, diverse HDL-associated proteins have been identified. Their functional categories include proteolysis, complement, coagulation, and lipid transport, among others, where they are thought to be involved in biological functions.2)
Several studies have shown that HDL function is disturbed under pathological conditions, and a remodeled HDL proteome may contribute to the observed functional changes. For example, the HDL proteome in patients with coronary artery disease is altered and can activate a proapoptotic pathway in endothelial cells.3) In patients with end-stage renal disease, the proteome is remodeled to contain more proteins belonging to acute phase reactants or the immune response.4) Recently, researchers have investigated the structure-function relationship of HDL in more detail. Particularly, HDL proteins including apolipoprotein A1 (apoA1) were shown to be involved in various biological functions.5) Riwanto et al.3) demonstrated that apoJ promoted an antiapoptotic pathway, while apoC3 activated the opposite signaling pathway in endothelial cells. In addition, apoL1, paraoxonase 1, and paraoxonase 3, carried by HDL3c, were reported to have anti-oxidant effects.6)
An optimized method for assessing HDL function has not been established. Cholesterol efflux capacity is the best-known index for HDL function, while endothelial nitric oxide (NO) production and vascular cell adhesion molecule (VCAM)-1 expression are often measured to evaluate the anti-inflammatory functions of HDL.7) Semi-quantitative proteomic analysis has been used to evaluate heterogeneous HDL particles.8) Thus, the aim of our study was to identify and rank HDL-associated proteins involved in representative HDL functions. We performed proteomic analysis to compare HDL-associated proteins in pooled samples according to cholesterol efflux capacity, NO production, and VCAM-1 expression by HDL.
METHODS
Study participants
Forty-eight patients with diverse cardiovascular risks were recruited for this study. Diverse health conditions may influence the composition and functions of HDL, and differences between HDL are needed for appropriate analysis of our study. Therefore, we included patients with various cardiovascular risk factors, unless they were in uncontrolled status (Supplementary Data 1). The study protocol was approved by the Institutional Review Board of Severance Hospital, Seoul, Korea (number 4-2013-0281). The investigation was conducted according to the principles stated in the Declaration of Helsinki. All participants provided written, informed consent. All patients enrolled in the study were statin-naïve or had discontinued prior lipid lowering agents for at least 3 months. Only patients without history of cardiovascular disease were eligible for wash-out of prior agents. Patients enrolled in this study and had coronary artery disease were all diagnosed for the first time and statin-naïve. Exclusion criteria are described in the Supplementary Data 1. At the time of enrollment, the patients were interviewed to obtain medical histories, followed by laboratory assessment.
Blood sampling and isolation of high-density lipoprotein
The patients were instructed to fast and avoid drinking alcoholic beverages or smoking for ≥12 hours before the collection of blood samples, which were analyzed within 4 hours. Lipid levels were analyzed using an auto-analyzer at a local laboratory certified by the Korean Society of Laboratory Medicine. HDL was isolated by ultracentrifugation as described in the Supplementary Data 1.
Tests of high-density lipoprotein function
Measurement of cholesterol efflux capacity, NO production, and VCAM-1 expression is described in the Supplementary Data 1.
Sample preparation
For proteomic analysis, HDL samples were pooled into equal amounts according to the highest and lowest quartiles of cholesterol efflux capacity, NO production, and VCAM-1 levels. Before mass spectral analysis, 80 μg of each pooled HDL sample was separated based on molecular weight in a NuPAGE gel (4–12% Bis-Tris gel; Invitrogen, Carlsbad, CA, USA). Methods for sample preparation and in-gel digestion are described in the Supplementary Data 1.
Mass spectral analysis
Ten microliters of each sample were injected into a reversed-phase Magic C18AQ column (15 cm × 75 µm) on an Eksigent MDLC system (Eksigent Technologies, Dublin, CA, USA). The nano HPLC system was coupled to an LTQ XL-Orbitrap mass spectrometer (Thermo Scientific, Waltham, MA, USA). Each liquid chromatography-mass spectrometry (MS)/MS file was analyzed using the SEQUEST algorithm in Proteome Discoverer 1.4 (Thermo Scientific). MS and MS/MS data were searched against the SwissProt human database (August 2016) with 20121. Detailed methods used for mass spectral analysis are available in the Supplementary Data 1.
Label-free quantitative analysis
The relative abundances of the proteins identified in pooled HDL samples showing the highest and lowest quartile values of cholesterol efflux capacity, NO production, and VCAM-levels were calculated based on the normalized spectral abundance factor (NSAF). Spectral count data (number of MS/MS spectra matched to the protein) were first extracted using Proteome Discoverer 1.4 software and exported into MS-EXCEL. The NSAF for each protein was calculated as follows:

in which the total number of MS/MS spectra matching peptides from protein k (SpC) was divided by protein length (L) and then divided by the sum of SpC/L for all N proteins.
For statistical analysis of the dataset, natural logarithm of each NSAF value was calculated, followed by Student's t-test of ln(NSAF) values obtained from the three replicates of the highest and lowest quartile values of each parameter. For the proteins showing relative abundance ratio >2 depending on three functional properties of HDL, Benjamini and Hochberg method-based false discovery ratio (FDR)-adjusted p values were calculated. A p value <0.05 was considered statistically significant. Additional analytical methods are described in the Supplementary Data 1.
RESULTS
Characteristics of study subjects
The demographic data of the study subjects are presented in Table 1. Of the 48 patients, 34 (71%) were male, with patient ages ranging from 30 to 80 years. The mean high-density lipoprotein-cholesterol (HDL-C) level was 45.7±13.2 mg/dL, whereas the mean low-density lipoprotein-cholesterol (LDL-C) level was 127±32 mg/dL. Patients with the highest quartile value of cholesterol efflux were younger and showed higher total- and LDL-C levels than those with the lowest quartile value. The characteristics and lipid profiles of the patients with the highest and lowest quartile values for NO production and VCAM-1 expression were not significantly different. Test results of each functional parameter according to quartile groups are presented in Figure 1.
Table 1
Clinical characteristics of the study subjects


Identification of HDL proteins associated with cholesterol efflux capacity, nitric oxide production, and vascular cell adhesion molecule-1 expression
For proteomic analysis, 80 μg of each HDL sample pooled according to the highest and lowest quartile values of the functional parameters was separated based on molecular weight. In-gel tryptic digestion of the stained protein bands was performed to identify the proteins. Through mass spectral analysis of the extracted peptides from each pooled HDL sample, 213, 184, and 199 proteins were identified, as shown in the Venn diagrams in Supplementary Figure 1.
Differential abundance of high-density lipoprotein proteins associated with cholesterol efflux capacity
Label-free quantification based on the spectral count was performed to identify HDL-associated proteins that were differentially abundant in samples with high functional parameter values. Only proteins with p<0.05 are shown in Figures 2, 3, 4. Table 1 lists the proteins that were highly ranked according to their differential abundance (relative abundance ratios ≥2 or ≤0.5, p value <0.05 from Student's t-test) in HDL with the highest quartile for each parameter. The spectral count and relative abundance ratios of all differentially abundant proteins (p value <0.05 from Student's t-test) are listed in Supplementary Tables 1, 2, 3. The functional categories of the proteins were classified and are presented in Supplementary Tables 4, 5, 6.
Figure 2
Proteins ranked by their differential abundance in HDL with high cholesterol efflux capacity. The ranking was determined by the NSAF ratio, which is log2-transformed in the figure.
HDL = high-density lipoprotein; NSAF = normalized spectral abundance factor.
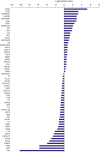
Figure 3
Proteins ranked by their differential abundance in HDL inducing high NO production. Ranking was determined by NSAF ratio, which is log2-transformed in the figure.
HDL = high-density lipoprotein; NO = nitric oxide; NSAF = normalized spectral abundance factor.

Figure 4
Proteins ranked by their differential abundance in HDL inducing high VCAM-1 expression levels. Ranking was determined by NSAF ratio, which is log2-transformed in the figure.
HDL = high-density lipoprotein; NSAF = normalized spectral abundance factor; VCAM = vascular cell adhesion molecule.

Twenty-seven proteins were more abundant and 33 were less abundant in HDL with the highest quartile compared to in HDL with the lowest quartile of cholesterol efflux capacity (Figure 2 and Supplementary Table 1). Fourteen proteins showed relative abundance ratios ≥2. However, when the p values were adjusted by multiple testing, only 6 proteins, thyroxine-binding globulin, alpha-1B-glycoprotein, plasma serine protease inhibitor, vitronectin, angiotensinogen, and serum amyloid A-4 (SAA4), met the criteria. In contrast, 14 proteins showed relative abundance ratios <0.5. After FDR adjustment by Benjamini and Hochberg method, only 3 proteins, complement C4-A, alpha-2-macroglobulin, and immunoglobulin mu chain C region, met the criteria (Table 1). When proteins that were more abundant in HDL with highest cholesterol efflux capacity were classified by functional categories, those belonging to “lipid metabolism” were most common. Among proteins less abundant in the same sample, those belonging to “immunity” were most common (Supplementary Table 4).
Differential abundance of high-density lipoprotein proteins associated with nitric oxide production
Nineteen proteins were significantly more abundant and seven were less abundant in HDL with the highest versus lowest quartile of NO production (Figure 3 and Supplementary Table 2). Four proteins showed relative abundance ratios ≥2. In contrast, no proteins showed relative abundance ratios of <0.5. When the p-values were adjusted by multiple testing, no proteins showed abundance ratios ≥2 or <0.5 (Table 2). The functional categories of proteins showing higher or lower abundance in HDL with high NO production varied considerably (Supplementary Table 5).
Table 2
Relative abundance ratios and functional categories of proteins highly ranked according to their differential abundance (NSAF ratio ≥2 or ≤0.5) in HDL with high cholesterol efflux, NO production, or VCAM-1 expression

For the proteins showing the relative abundance ratios ≥2 depending on three functional properties of HDL, the Benjamini and Hochberg method-based false discovery rate-adjusted p values were calculated.
HDL = high-density lipoprotein; NO = nitric oxide; NSAF = normalized spectral abundance factor; VCAM = vascular cell adhesion molecule.
Differential abundance of high-density lipoprotein proteins associated with vascular cell adhesion molecule-1 expression
Fifteen proteins were significantly more abundant and nine were less abundant in HDL with the highest quartile than in HDL with the lowest quartile of VCAM-1 expression (Figure 4 and Supplementary Table 3). Three proteins showed relative abundance ratios ≥2. Six proteins showed relative abundance ratios <0.5. After adjusting the p values, no proteins showed abundance ratios of ≥2 or <0.5 (Table 1). When the proteins that were more abundant in HDL showing high VCAM-1 expression were classified by functional categories, those belong to “immunity” were most common (Supplementary Table 6).
DISCUSSION
In the present study, we identified and ranked HDL proteins associated with three functional parameters by comparing HDL samples with the highest and lowest values. Proteins belonging to diverse functional categories, including lipid metabolism and immunity, showed different abundances in HDL with high cholesterol efflux or anti-inflammatory effects. Notably, we identified several protein-function relationships associated with the effects of HDL that have not been reported previously. For example, after adjustment of the p-values, significant associations were observed between nine proteins and cholesterol efflux.
Researchers have reported relationship between abundance of several HDL-associated proteins and cholesterol efflux capacity. Gordon et al.9) and Pamir et al.10) identified several porteins such as apoA1, Gelsolin, antithrombin-III, apoE, and plasminogen as determinants of this capacity. Some proteins including complement C39) or apoC311) showed negative relationships to cholesterol efflux capacity. However, the relationships have not been consistent among all studies.12)
Vitronectin is known to play a role in proteolysis, hemostasis, and complement regulation.13)14) In the present study, this protein was found to be associated with high cholesterol efflux capacity, with a relative abundance ratio of 3.29. Vitronectin has been reported to be more enriched in HDL from the control group than from patients with coronary artery disease.15) However, the association between cholesterol efflux and vitronectin has not been demonstrated previously. Further mechanistic investigations are required to understand the biological role of HDL-associated vitronectin.
Our data revealed a positive association between SAA4 and cholesterol efflux. SAA4 is known to be present in HDL.15) SAA has been reported to induce cellular lipid release and HDL generation, both in an ATP-binding cassette transporter A1 (ABCA1)-dependent and ABCA1-independent manner.16)17) In addition, SAA was suggested to promote cholesterol efflux via SR-B1.18) Our results provide insight into the association between SAA abundance and cholesterol efflux. However, this protein is also known to have a proinflammatory effect.19)
Several studies have examined the proteins evaluated in our study, providing insights into their roles. We observed that insulin-like growth factor-binding protein 3 (IGFBP3) in HDL was marginally associated with high cholesterol efflux capacity. Many, but not all, previous studies showed that IGFBP3 levels are positively correlated with blood HDL-C levels.20)21) However, the relationship between IGFBP3 and cholesterol efflux capacity has not been reported. In contrast, IGFBP3 was reported to decrease inflammation by inhibiting the JNK and nuclear factor-κB pathways.22) In addition, IGFBP3 attenuates tumor necrosis factor-α-mediated apoptosis in microvascular endothelial cells.23) However, a previous study showed that blood IGFBP3 levels are correlated with carotid intima-media thickness24); thus, the role of this protein in atherosclerosis requires clarification. Originally identified as a neuroprotective factor, pigment epithelium-derived factor (PEDF) influences angiogenesis and apoptosis in multiple cell types.25) Blood PEDF levels, which can be elevated by a counter-regulatory response, were associated with vascular inflammation and carotid intima-media thickness.26) Interestingly, PEDF was reported to inhibit endothelial activation by suppressing the generation of reactive oxygen species.27) In our study, PEDF was weakly related to cholesterol efflux capacity, suggesting that this protein is involved in vascular protection.
In this study, apoA1, the most abundant protein in HDL, was not ranked highly as a differentially abundant protein for HDL functions. Although the level of this protein is much greater than that of other high-ranked proteins, its lack of functional associations may explain the smaller difference between apoA1 levels in HDLs with the highest and lowest test values. ApoA4 was 1.44-fold more abundant in the highest quartile group of cholesterol efflux capacity, while other apoAs did not show difference according to cholesterol efflux capacity groups either. With regard to NO production and VCAM-1 expression, no apoAs revealed significant difference depending on the functional property of HDL.
This study had some limitations. First, clinical risk factors could not be adjusted during the analysis. However, most risk factors were similar between sample groups, except for marginal differences in age and cholesterol levels. Second, elucidation of the action mechanism of proteins was beyond the scope of our study. For instance, for some proteins, such as thyroxine-binding globulin, alpha-1B-glycoprotein, plasma serine protease inhibitor, and angiotensinogen, a role in cholesterol efflux or inflammation remains to be suggested. Further studies are needed to confirm the roles of these proteins. Third, we did not analyze lipid component of HDL. However, the focus of our study was proteomic analysis, and we obtained appropriate results. Because lipid composition can be associated with biological function of HDL,28) lipidomic analysis might have added further information to our results. Fourth, we did not separately analyze subfractions of HDL. Although comparison of subfractions, such as HDL2 or HDL3, could provide some additional data, it was not necessary for the purpose of our study that was to investigate associations between multiple HDL proteins and parameters of HDL function. In previous studies, researchers used total HDL3)7) or HDL subfractions6) according to their aims.
In summary, in this study, proteins showing greater or lower abundance in HDLs with high functional capacities were identified and ranked. Nine HDL proteins were newly identified as being linked to the cholesterol efflux capacity of HDL. Proteins abundant in HDL with high functional parameters belonged to diverse biological categories.
 XML Download
XML Download