

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
A heterogeneous lung inflammation in asthma is characterized by airway remodeling, airway hyperresponsiveness (AHR), and mucus hypersecretion, involving various immune cells and mediators.1 Asthma has long been regarded as a T-helper (Th) 2-dependent, immunoglobulin (Ig) E-mediated disease. However, emerging evidence suggests that asthma is a complex disease displaying various endotypes: eosinophilic, neutrophilic, mixed eosinophilic/neutrophilic, and paucigranulocytic23 or Th2-predominant, Th2/Th17-predominant, and Th2/Th17-low asthma.45
Approximately 358 million people suffer from asthma worldwide and its incidence continues to increase.6 Current guidelines for asthma management rely on various agents, including new and emerging drugs; however, the medical need is still unmet. Asthma is a consequence of complex host-environment interactions and has many phenotypic features involving various immune responses.7 Thus, there is a continuous research effort for better targeted and personalized therapy as well as a deeper understanding of the complex pathophysiology of this disease.8
A variety of animal models of asthma have been established not only for understanding the pathophysiology, but also for evaluating the efficacy and safety of new therapeutic agents.9 Mice are the most widely used model animals because of the availability of comprehensive genetic information and transgenic, congenic, and knockout animals, which allow examining the effect of a specific gene in disease onset and development. Other advantages are that numerous mouse inbred strains are available and methods for measuring cytokines, growth factors, and cell-surface molecules are well established.910 Mice do not naturally develop human-like asthma; thus, to investigate the underlying processes, an asthmatic-like reaction, mainly representing Th2-type response-mediated disease, has been induced using various allergens.10
There are various sensitization and challenge protocols to induce experimental asthma in mice. Chicken egg ovalbumin (OVA) is the most frequently used allergen to induce a Th2-skewed response because of its high purity and immunodominant epitopes. Moreover, using OVA enables immunological application, for example, in transgenic mice bearing T cells expressing OVA-specific T-cell receptors.11 Generally, mice are first systemically immunized with OVA, usually by intraperitoneal (i.p.) injection, followed by repetitive inhalation of OVA by intranasal (i.n.),121314 intratracheal (i.t.),151617 or nebulized formulation (aerosol),131819 resulting in a robust influx of inflammatory cells in the lungs and elevation of IgE levels. However, sensitizing mice with OVA alone via inhalation induces immune tolerance rather than overt asthma-like airway responses.20 Therefore, a refined murine model that more accurately and strongly induces asthma-related immune responses is needed.
Moreover, the conventional methods for establishing murine asthma models have several limitations. For example, mice have to be anesthetized for repetitive allergen inhalation via the i.n. route. Repeated anesthesia for consecutive days can be stressful for the mice as well as the researchers, and anesthesia itself is an artificially controlled condition and can result in variable outcomes. Further, the amount of inhaled allergen can differ among individuals, causing phenotypic variation even within an experimental group. Moreover, the conventional models do not reflect heterogeneous human asthma because of lack of non-allergic asthmatic responses accompanied by Th17 and neutrophils.21
Dendritic cells (DCs) are the most specialized antigen-presenting cells that play a critical role in initiating and directing asthma.722 Once activated, mature DCs migrate to local lymph nodes where they interact with naïve T cells to drive the differentiation of T cells into Th1, Th2, Th9, Th17, or regulatory T cells.22 DCs are important targets for studying the pathophysiology and treatment of asthma because they play a crucial role in striking a balance between immune activation and immune tolerance.232425
To address the above issues, in the present study, we aimed to develop a reliable murine model of asthma through direct i.v. administration of OVA-loaded DCs into the mice, thereby circumventing the need for anesthesia. This method induces more severe inflammation than conventional methods and allows for high productivity. In addition, we examined the immunologic effect of direct transfer of DCs and checked the variability between individuals.
MATERIALS AND METHODS
Mice
Female C57BL/6J mice, 4-5 weeks of age, were purchased from Japan SLC, Inc. (Shizuoka, Japan) and were maintained under specific pathogen-free conditions. The use of animals in this study was approved by the Ethics Committee and Institutional Animal Care and Use Committee of Yonsei University Health System (2017-0342), and the study was conducted according to the International Association of Veterinary Editors guidelines on animal experiments.
Murine models of asthma
To establish a conventional asthma model, a previously published protocol was used,12 with slight modification. Mice were sensitized on days 0 and 14 via i.p. injection of 50 μg OVA (grade V; Sigma-Aldrich, St. Louis, MO, USA) emulsified in 1.32 mg alum gel (Sigma-Aldrich) in a total volume of 200 μL. Phosphate-buffered saline (PBS)-treated mice served as the negative control group. Anesthetized mice were challenged i.n. with 150 μg of OVA in 30 μL of PBS for 5 consecutive days starting from day 21. Mice were analyzed 24 hours after the last challenge (Fig. 1A).
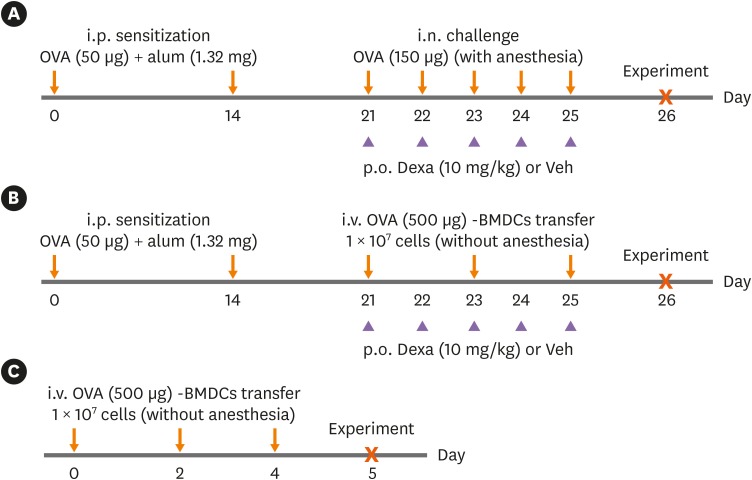
Fig. 1
Experimental protocol for the development of asthma in mice. (A) Conventional OVA-induced asthma model by i.p. OVA sensitization and i.n. OVA challenge under anesthesia. (B) Asthma model established by i.v. transfer of DCs. After i.p. OVA sensitization, OVA-BMDCs were injected i.v. instead of i.n. (C) I.v. transfer of OVA-BMDCs alone without i.p. OVA sensitization was included in some experiments. There was no need for anesthesia for i.v. transfer of DCs. For some experiments, mice of asthma group were treated with vehicle or dexamethasone during the period of i.n. OVA challenge or i.v. transfer of DCs.
OVA, ovalbumin; BMDC, bone marrow-derived dendritic cell; OVA-BMDC, ovalbumin-loaded bone marrow-derived dendritic cell; DC, dendritic cell; i.p., intraperitoneal; i.n., intranasal; i.v., intravenous; Dexa, dexamethasone; Veh, vehicle.
![]()
The murine model of asthma was established via transfer of OVA-loaded DCs. On days 0 and 14, i.p., OVA sensitization was done as per the conventional method. On days 21, 23 and 25, 1 × 107 OVA (500 μg)-loaded bone marrow-derived DCs (BMDCs) (OVA-BMDCs) in a total volume of 200 μL were injected i.v. (via the tail vein) into non-anesthetized mice. Mice were analyzed 24 hours after the last transfer (Fig. 1B). In addition, i.v. OVA-BMDC transfer without i.p. sensitization was used (Fig. 1C).
In some experiments, dexamethasone (10 mg/kg; Sigma-Aldrich) or vehicle (0.25% PEG400, 0.25% Tween20 in dH2O) was administered to the mice by oral gavage 2 hours prior to i.n. challenge or i.v. transfer for 5 consecutive days starting from day 21.
Generation of OVA-BMDCs
BMDCs were generated from BM using a conventional method.26 Whole BM cells isolated from C57BL/6J mice were cultured in RPMI 1640 medium supplemented with 100 units/mL of penicillin/streptomycin (Lonza, Basel, Switzerland), 10% fetal bovine serum (Lonza), 50 μM mercaptoethanol (Lonza), and 20 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF) plus 5 ng/mL interleukin (IL)-4 at 37°C in the presence of 5% CO2. On days 3 and 6, the medium was refreshed. On day 8, BMDCs were harvested and incubated in 24-well multiwell plates (1 × 106 cells/well) in the presence of OVA protein (500 μg/mL) for 24 hours. On day 9, OVA-BMDCs were harvested.
Histological assessment of lung tissue
Lung tissue was fixed in 10% neutral buffered formalin and embedded in paraffin. Paraffin-embedded tissue sections were stained with hematoxylin and eosin (H&E) or periodic acid-Schiff (PAS) solution. Lung inflammation and goblet cell hyperplasia were graded using a previously reported semiquantitative scoring system.27 To grade inflammatory cell infiltration, peribronchial cell counts were determined on the basis of a 5-point grading system as follows: 0, normal; 1, few cells; 2, a ring of inflammatory cells one cell layer deep; 3, a ring of inflammatory cells of 2–4 cells deep; and 4, a ring of inflammatory cells of more than 4 cells deep. Eight fields were counted for each section, and the mean score from 5–7 mice per group was calculated. To quantify airway goblet cells, the following 5-point grading system was used: 0, < 0.5% PAS-positive cells; 1, < 25%; 2, 25%–50%; 3, 50%–75%; and 4, > 75%. Eight fields were counted for each section, and the mean area or mean score from 5–7 mice per group was calculated. Analyses were conducted in a blinded manner, and slides were presented in random order for each examination.
Analysis of AHR
AHR was measured by whole-body plethysmography (Buxco, Wilmington, NC, USA) with a technical support from the Korea Mouse Phenotyping Center (Seoul, Korea) as previously described.28 AHR was represented as enhanced pause (Penh) in response to increasing concentrations of methacholine (6.25–50 mg/mL). Penh values measured during this period were averaged and expressed as absolute Penh values.
Flow-cytometric analysis and intracellular cytokine staining
To analyze myeloid cells, single-cell suspensions were blocked with Fc Block (anti-CD16/32; eBioscience, San Diego, CA, USA) at 4°C for 15 minutes and then stained with fluorochrome-conjugated anti-CD3e, anti-CD11b, anti-CD103, anti-F4/80, anti-MHCII (eBioscience), anti-CD4, anti-Ly6G, anti-Siglec-F (BD Biosciences, San Jose, CA, USA), and anti-CD11c, anti-CD19, and anti-CD64 (BioLegend, San Diego, CA, USA) antibodies at 4°C for 30 minutes. Cells stained with appropriate isotype-matched antibodies were used as negative controls. Cells were fixed using an IC fixation buffer (eBioscience) in accordance with the manufacturer's instructions. The stained cells were analyzed using a BD LSR II Fortessa flow cytometer (BD Biosciences) and FlowJo software (Tree Star, Inc., Ashland, OR, USA). T-cell transcription factors and Treg marker expression were analyzed by staining with fluorochrome-conjugated anti-CD3e (eBioscience), anti-CD4, and anti-CD25 (BD Biosciences) antibodies, followed by permeabilization with a fixation/permeabilization kit (eBioscience) in accordance with the manufacturer's instructions and intracellular staining with anti-T-bet, anti-GATA3, anti-RORγt and anti-Foxp3 (eBioscience) antibodies. To restimulate cells with the antigen, lung cells (1 × 106 cells/well) and spleen cells (2 × 106 cells/well) were restimulated with OVA protein (10 μg/mL) at 37°C. Intracellular cytokines of CD4+ T cells were quantified after 2 hours of incubation with the antigen, 4 hours of incubation with GolgiStop (eBioscience), and permeabilization, followed by intracellular staining with fluorochrome-conjugated anti-interferon (IFN)-γ, anti-IL-4, anti-IL-5 and anti-IL-17 (eBioscience) antibodies. To determine cytokine levels by an ex vivo recall assay, supernatants were harvested after 72 hours of incubation with the antigen, and cytokine production was analyzed via enzyme-linked immunosorbent assay (ELISA).
ELISA of antigen-specific antibodies
Serum antigen-specific antibodies were quantified by sandwich ELISA as previously described,29 with minor modifications. Plates were coated with OVA (1 mg/mL). Serum dilutions were 1:50 for OVA-specific IgE and 1:10,000 for OVA-specific IgG1. Biotinylated anti-mouse IgE (clone: R35-118) and anti-mouse IgG1 (clone: A85-1) were used, followed by incubation with horseradish peroxidase-conjugated streptavidin.
Statistical analysis
All data are representative of at least 3 independent experiments with consistent results, with 5–7 mice per group in each experiment. Means were compared using 1-way analysis of variance followed by Tukey's multiple comparison test when data were normally distributed using Prism 5 (version 5; GraphPad Software, San Diego, CA, USA). To assess normality, the Shapiro-Wilk test was used. A P value < 0.05 was considered significant.
RESULTS
Mice receiving i.v. OVA-BMDCs exhibit more severe lung inflammation than conventional asthma model mice
To evaluate the asthma phenotype, we first analyzed lung histopathology. H&E staining revealed increased peribronchial, perivascular, and parenchymal infiltration of inflammatory cells in conventional asthma model mice compared to negative control mice. I.v. OVA-BMDC-transferred asthma model mice developed stronger inflammatory cell infiltration than did conventional model mice. Interestingly, mice receiving i.v. OVA-BMDCs alone, without i.p. OVA challenge, also showed inflammatory cell infiltration (Fig. 2A).
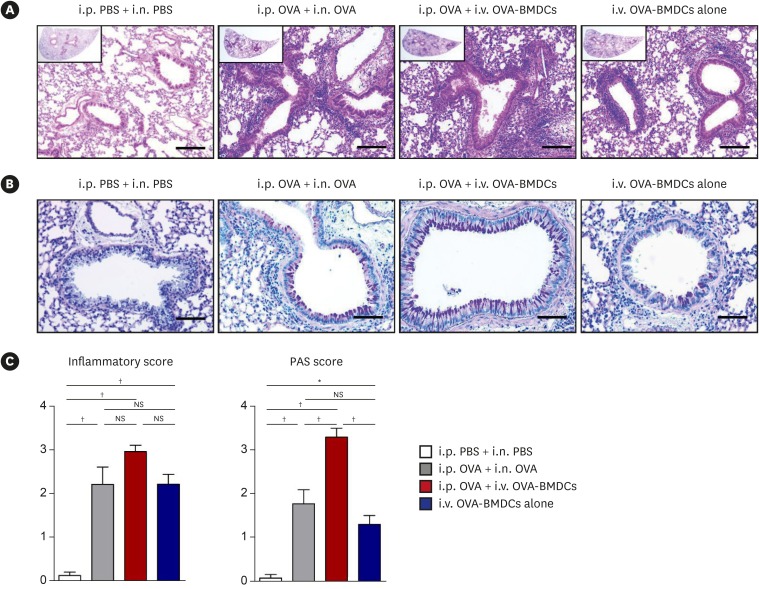
Fig. 2
Assessment of lung histology in the various asthma models. (A) Representative H&E-stained lung tissue sections (original magnification: 100×, magnified image of the 10× image in the left upper quadrant, scale bar: 100 μm) and (B) Representative PAS-stained sections (original magnification: 200×, scale bar: 50 μm) in control mice (i.p. PBS and i.n. PBS-challenged), conventional asthma model mice (i.p. OVA-sensitized and i.n. OVA-challenged), alternative asthma model mice (i.p. OVA-sensitized and i.v. injected with OVA-BMDCs [1 × 107 cells]), and mice injected i.v. with OVA-BMDCs (1 × 107 cells) alone, without i.p. OVA sensitization. (C) Lung inflammation and PAS-positive cell score. Figures comprise histological examination data representative of 5 to 7 mice per group. All data are representative of 3 independent experiments with similar results. Data are expressed as the mean ± standard deviation. Means were compared using 1-way analysis of variance followed by Tukey's multiple comparison test when data were normally distributed.
PBS, phosphate-buffered saline; OVA, ovalbumin; BMDC, bone marrow-derived dendritic cell; OVA-BMDC, ovalbumin-loaded bone marrow-derived dendritic cell; i.p., intraperitoneal; i.n., intranasal; i.v., intravenous; H&E, hematoxylin and eosin; PAS, periodic acid–Schiff; NS, not significant.
*P < 0.01, †P < 0.001.
![]()
PAS staining revealed that the goblet cell population was expanded in conventional asthma model mice compared to negative control mice. Strikingly, this population was the most increased in i.v. OVA-BMDC-transferred asthma model mice (Fig. 2B).
These microscopic findings were confirmed by semiquantitative scoring of the inflammatory cell infiltration and PAS-positive cells. Notably, there was less variation in inflammatory scores and PAS scores among individuals in the i.v. OVA-BMDC-transferred than in the conventional model group (mean ± standard deviation: 2.97 ± 0.30 vs. 2.22 ± 0.80 and 3.32 ± 0.41 vs. 1.79 ± 0.65, respectively; Fig. 2C).
In addition, we checked the phenotype of i.p. PBS-sensitized and i.v. PBS-BMDCs-transferred mice, because they are an ideal negative control group for the i.p. OVA-sensitized and i.v. OVA-BMDC-transferred asthma group. In i.p. PBS-sensitized and i.v. PBS-BMDCs-transferred mice, the infiltration of inflammatory cells and goblet cells (Supplementary Fig. S1A-C) and OVA-specific IgE and IgG1 (Supplementary Fig. S1D) was not increased compared to the asthma groups; this was similar to the results in i.p. PBS-sensitized and i.n. PBS-challenged mice. Thus, in subsequent experiments, we used i.p. PBS-sensitized and i.n. PBS-challenged mice as negative control group.
The severity of asthma correlates with the number of OVA-BMDCs transferred
To elucidate the effect of the number of BMDCs injected in i.v. OVA-BMDC-transferred asthma model mice, we investigated lung histology in mice injected with increasing numbers of OVA-BMDCs (2 × 106, 5 × 106 and 1 × 107 cells). At a dose of 2 × 106 cells, inflammatory cell infiltration was comparable or slightly higher than that in conventional asthma model mice. The inflammation increased gradually as the cell number increased (Supplementary Fig. S2A). Goblet cell infiltration at 2 × 106 OVA-BMDCs reached a level similar to that in conventional model mice and then increased with increasing cell number, reaching a maximum at 1 × 107 cells (Supplementary Fig. S2B). These findings were validated by inflammatory cell infiltration and PAS score findings. As shown in Supplementary Fig. S2C, i.p. OVA challenge combined with i.v. 1 × 107 OVA-BMDCs induced the most severe lung inflammation and goblet cell infiltration.
Next, we aimed to find the appropriate concentration of OVA for priming BMDCs. CD11c and MHCII double-positive DCs were examined. BMDCs were stimulated with increasing doses of OVA (0, 10, 100, and 500 μg/mL) (Supplementary Fig. S3A and B). The expression of CD80 and CD86, co-stimulatory molecules of DCs, increased with increasing OVA concentration. Expression of OX40L, a Th2 inducer, also increased with increasing OVA concentration (Supplementary Fig. S3C). Based on the data, 500 μg/mL OVA for priming BMDCs was considered effective for inducing asthma in the present study.
Mice receiving i.v. OVA-BMDCs present stronger systemic and local allergy-associated immune responses than conventional asthma model mice
AHR determined by methacholine exposure, a key feature of allergic asthma, was increased in conventional asthma model mice than in negative control mice. I.v. OVA-BMDC-transferred asthma model mice also enhanced AHR when accompanied by i.p. OVA sensitization, comparable to conventional asthma model mice (Fig. 3A).
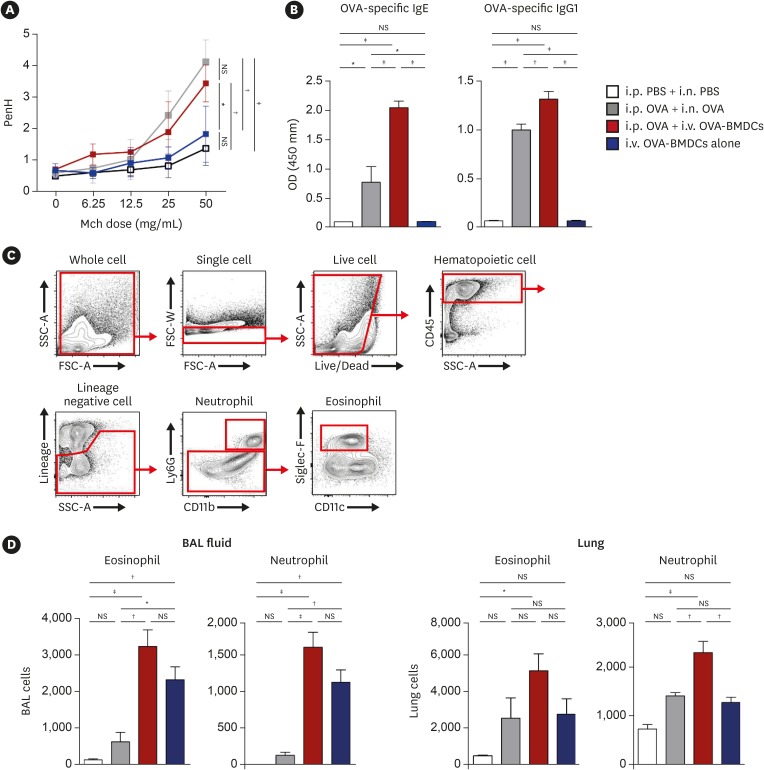
Fig. 3
Evaluation of AHR, serum OVA-specific Ig, and myeloid cells. (A) AHR represented as Penh in response to methacholine. (B) Serum OVA-specific Ig measured by ELISA. (C) Gating strategy for the examination of myeloid cells in BAL fluid and lungs. (D) Populations of eosinophils and neutrophils in BAL fluid and lung. All data are representative of 3 independent experiments with similar results. Data are expressed as means ± standard deviation. Means were compared using 1-way analysis of variance followed by Tukey's multiple comparison test when data were normally distributed.
AHR, airway hyperresponsiveness; PBS, phosphate-buffered saline; OVA, ovalbumin; BMDC, bone marrow-derived dendritic cell; OVA-BMDC, ovalbumin-loaded bone marrow-derived dendritic cell; i.p., intraperitoneal; i.n., intranasal; i.v., intravenous; Mch, methacholine; Ig, immunoglobulin; BAL, bronchoalveolar lavage; ELISA, enzyme-linked immunosorbent assay; NS, not significant; OD, optical density.
*P < 0.05, †P < 0.01, ‡P < 0.001.
![]()
The serum levels of OVA-specific IgE and IgG1 were elevated in conventional asthma model mice when compared to negative control mice, but were more strongly increased in i.v. OVA-BMDC-transferred asthma model mice. Particularly, a more than 2-fold increase in OVA-specific IgE was seen in the i.v. OVA-BMDC-transferred asthma model mice, with less variation among individuals than in the conventional asthma model mice (2.04 ± 0.28 vs. 0.77 ± 0.60) (Fig. 3B).
Next, we examined the population of eosinophils and neutrophils in bronchoalveolar lavage (BAL) fluid and the lungs (Fig. 3C). In BAL fluid, eosinophil and neutrophil counts were significantly higher in i.v. OVA-BMDC-transferred asthma model mice, independent of prior i.p. OVA sensitization, than in conventional asthma model mice. In the lungs, eosinophil and neutrophil infiltration was also higher in the i.v. OVA-BMDC-transferred than in the conventional asthma model mice. Interestingly, i.v. transfer of OVA-BMDCs alone, without i.p. OVA sensitization, was sufficient to induce eosinophil and neutrophil recruitment (Fig. 3D).
Together, these results suggested that systemic (serum IgE and IgG1) as well as local (eosinophils and neutrophils in the lungs and bronchus) immune responses were robustly induced in i.v. OVA-BMDC-transferred asthma model mice. In addition, i.p. OVA sensitization is required for inducing asthma-related systemic immune responses.
Mice receiving i.v. OVA-BMDCs display an aggravated Th2/Th17-mediated immune response
To examine the T-cell immune response, we measured cytokine expression in OVA-restimulated lungs, spleen and mediastinal lymph node (MLN) cells. The expression of the Th2 cytokines IL-5 and IL-13 in the lungs was the highest in i.v. OVA-BMDC-transferred asthma model mice, followed by mice receiving i.v. OVA-BMDCs alone (Fig. 4A). A similar expression pattern was noted in the spleen and MLN cells (Supplementary Fig. S4A and B). Meanwhile, the levels of IFN-γ and IL-17 in the lungs, spleen, and MLN cells were elevated in the i.v. OVA-BMDC-transferred asthma model mice, but were the highest in the mice receiving i.v. OVA-BMDCs alone; the exception being IL-17 in spleen cells.
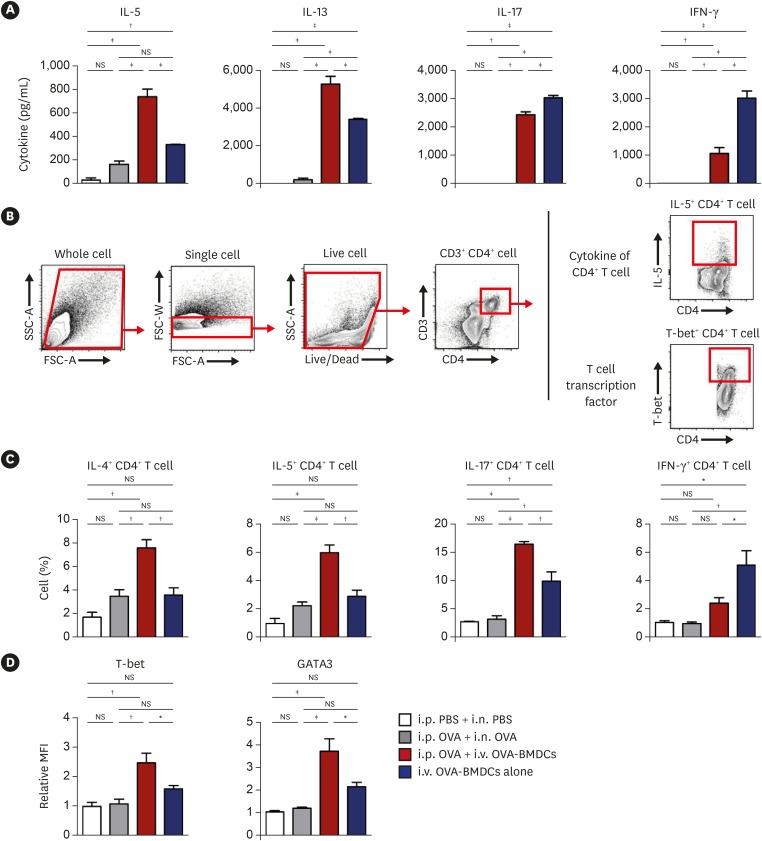
Fig. 4
Cytokine profiles and analysis of T cell transcription factors in CD4+ T cells of lung. (A) Cytokines were measured by ex-vivo recall assays with OVA protein (10 μg/mL) in isolated lung cells. (B) Gating strategy for the examination of intracellular cytokines and transcription factors of CD4+ T cells. (C) Intracellular levels of IL-4, IL-5, IL-17 and IFN-γ as measured by flow cytometry after stimulation with OVA (10 μg/mL) for 2 hours with GolgiStop. (D) T-cell transcription factors showing relative MFI of T-bet and GATA3. All data are representative of 3 independent experiments with similar results. Data are expressed as the mean ± standard deviation. Means were compared using one-way analysis of variance followed by Tukey's multiple comparison test when data were normally distributed.
PBS, phosphate-buffered saline; OVA, ovalbumin; BMDC, bone marrow-derived dendritic cell; OVA-BMDC, ovalbumin-loaded bone marrow-derived dendritic cell; i.p., intraperitoneal; i.n., intranasal; i.v., intravenous; IL, interleukin; IFN, interferon; MFI, mean fluorescence intensity; NS, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.
![]()
Next, we checked the expression of intracellular cytokines in CD4+ T cells of the lung (Fig. 4B). The populations of IL-4+ CD4+ and IL-5+ CD4+ T cells were elevated in conventional asthma model mice and in mice receiving i.v. OVA-BMDCs alone, but were more strongly increased in i.v. OVA-BMDC-transferred asthma model mice. The population of IL-17+ CD4+ T cells was the largest in i.v. OVA-BMDC-transferred asthma model mice, whereas the population of IFN-γ+ CD4+ T cells was the largest in mice receiving i.v. OVA-BMDCs alone (Fig. 4C).
To confirm T-cell polarization, we examined T-cell transcription factors of the lung. The relative mean fluorescence intensity (MFI) was measured in each group compared to that in negative control mice. The levels of T-bet and GATA3 were comparable between the negative control and conventional asthma model mice. However, in i.v. OVA-BMDC-transferred asthma model mice, the level of T-bet was elevated by more than 2-fold, whereas a more than 3-fold increase was observed in the level of GATA3, suggesting that the Th2 immune response was robustly induced especially in these mice (Fig. 4D).
Collectively, these data suggested that i.v. transfer of OVA-BMDCs induced a stronger T-cell immune response than the conventional method. Th1, Th2 and Th17 levels were all augmented, but i.p. OVA sensitization before i.v. injection of OVA-BMDCs induced a stronger Th2/Th17 immune response, representing heterogeneous human asthma.
Mice receiving i.v. OVA-BMDCs are partially resistant to steroid treatment
In addition, we evaluated the response to therapeutic application of dexamethasone, since resistance to steroid treatment is another major issue in severe human asthma. In conventional asthma model mice, infiltration of inflammatory cells (Fig. 5A) and hyperplasia of goblet cell (Fig. 5B) were significantly decreased after dexamethasone administration. However, i.v. OVA-BMDC-transferred asthma model mice showed partially decreased populations of inflammatory (Fig. 5A) and goblet cells (Fig. 5B) after the same dexamethasone treatment.
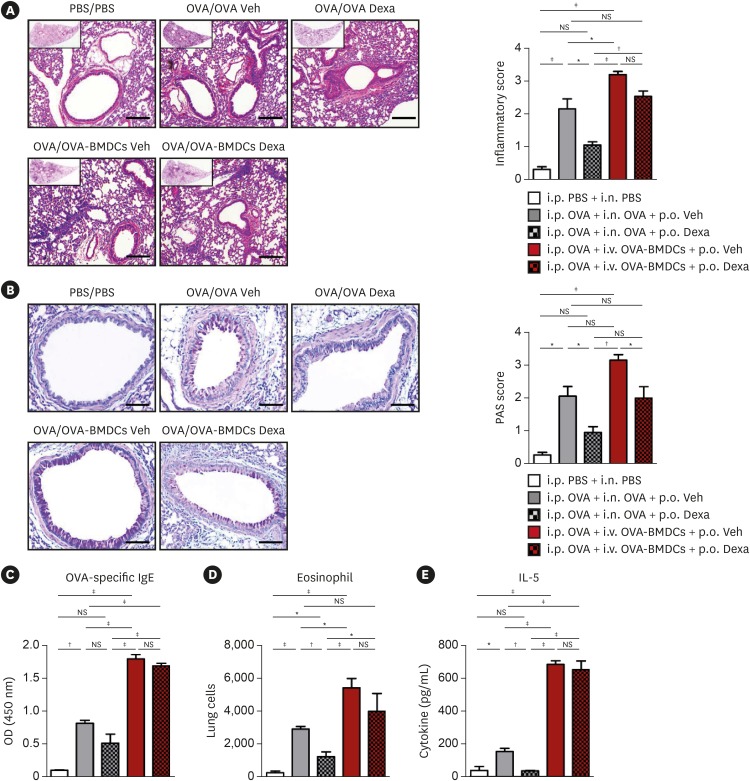
Fig. 5
Effect of dexamethasone administration on lung histology and immune responses. (A) Representative H&E-stained lung tissue sections (original magnification: 100×, magnified image of the 10× image in the left upper quadrant, scale bar: 100 μm) with lung inflammation score and (B) Representative PAS-stained sections (original magnification: 200×, scale bar: 50 μm) with PAS-positive cell score in control i.p. PBS and i.n. PBS-challenged mice (PBS/PBS), conventional i.p. OVA-sensitized and i.n. OVA-challenged asthma model mice (OVA/OVA), new i.p. OVA-sensitized and i.v. transfer of OVA-BMDCs asthma model mice (OVA/OVA-BMDCs). Asthma mice were administered dexamethasone (10 mg/kg) or vehicle. Figures show the histological examination data representative of 5 to 7 mice per group. (C) Serum OVA-specific IgE measured by ELISA. (D) Population of eosinophils in lung. (E) Cytokines measured by ex-vivo recall assays with OVA protein (10 μg/mL) in isolated lung cells. Data are expressed as mean ± standard deviation. Means were compared using 1-way analysis of variance followed by Tukey’s multiple comparison test when data were normally distributed.
PBS, phosphate-buffered saline; OVA, ovalbumin; BMDC, bone marrow-derived dendritic cell; OVA-BMDC, ovalbumin-loaded bone marrow-derived dendritic cell; Dexa, dexamethasone; Veh, vehicle; i.p., intraperitoneal; i.n., intranasal; i.v., intravenous; H&E, hematoxylin and eosin; PAS, periodic acid–Schiff; IL, interleukin; ELISA, enzyme-linked immunosorbent assay; NS, not significant; OD, optical density; IgE, immunoglobulin E.
*P < 0.05, †P < 0.01, ‡P < 0.001.
![]()
The serum level of OVA-specific IgE was also decreased in conventional asthma model mice after dexamethasone administration, whereas it was partially attenuated in i.v. OVA-BMDC-transferred asthma model mice (Fig. 5C). Moreover, dexamethasone treatment significantly reduced eosinophil population (Fig. 5D) and IL-13 expression in the lung (Fig. 5E) of the conventional asthma model, whereas it had limited effect in i.v. OVA-BMDC-transferred asthma model mice.
These results suggested i.v. OVA-BMDC-transferred asthma model mice are partially resistant to steroid treatment, similar to that in severe human asthma.
DISCUSSION
In the present study, we successfully developed a murine asthma model by i.v. transfer of OVA-BMDCs. The model showed typical histological (Fig. 2) and immunological alterations (Figs. 3 and 4) representing heterogeneous human-like asthma, with low variability among individual animals (Figs. 2C and 3B). In addition, this model was partially resistant to steroid treatment, a characteristic of severe human asthma (Fig. 5). To induce robust systemic immune responses, we used antigen-presenting DCs, which are pivotal in T-cell differentiation and in the initiation and maintenance of allergic responses. The allergen OVA was directly transferred into the mice with the DCs, resulting in a more severe asthma phenotype.
Several previous murine models of asthma relied on the transfer of DCs. For example, OVA-loaded splenic DCs were administered i.p. once for sensitization.30 However, the number of spleen cells available was insufficient to generate multiple murine models simultaneously. Therefore, studies have used BMDCs cultured for 6 to 8 days in medium containing GM-CSF. For example, OVA-BMDCs have been administered i.n.3132 or i.t.3334 to mice for sensitization. However, mice were anesthetized for OVA-BMDC administration in these studies. Furthermore, all of these previous models using DCs require repeated secondary i.n. or aerosol OVA challenge and thus, were not advantageous compared to conventional methods.
Our new asthma model provides significant research advantages compared to current models. First, our model better mimics heterogeneous human asthma, with a boosted immune response. In human asthma, Th2-mediated eosinophilic inflammation is the most common endotype, but Th1 and Th17 cells are also involved in the pathogenesis and are related with the heterogeneous features of asthma.452435 Raundhal et al.36 reported increased expression of IFN-γ+ CD4 (Th1) cells in patients with severe asthma. Th17 cells, which mediate neutrophil recruitment, are associated with resistance to steroid therapy and severe clinical course of asthma.37 Consequently, neutrophil infiltration is related with a severe disease phenotype in humans338 and mice.39 Patients with severe or steroid-resistant asthma show Th2 mixed with Th17 or Th1 immune responses.4041 Our model showed elevated Th17 immune responses as well as a Th2 response, including eosinophil and neutrophil infiltration, and was partially resistant to steroid treatment, thus being a good model of human asthma.
Secondly, this DCs-based asthma model could be useful for immunologic studies on the functions of DCs and T-cell responses in asthma. For example, manipulation of DCs in vitro, such as treatment with specific cytokines or adjuvants, followed by in vivo administration might aid in investigating the role of different pathways in DCs and elucidating the disease mechanism.31 Furthermore, in vivo transfer of allergen-loaded DCs is useful for investigating the pathogenesis of asthma while conserving the experimental animals. For example, DCs isolated from type I IFN receptor-deficient mice, which cannot initiate Th2 responses42 or from IL-33 knockout mice, which have alleviated asthma features,43 could be applied for immunological studies.
Thirdly, the degree of asthma inflammation can be controlled by varying the number of cells injected. More severe asthma could be established by increasing the cell number; 1 × 107 cells induced the most severe lung inflammation and goblet cell hyperplasia in the present study. This demonstrated that disease severity and asthma type can be adjusted to the research aims.
Fourthly, notably, asthma can be induced with less variation among individuals. In our data, inflammatory and PAS scores of lung histology (Fig. 2C) as well as serum OVA-specific IgE levels (Fig. 3B) were more constant in i.v. OVA-BMDC-transferred asthma model mice than in conventional asthma model mice. This advantage is in contrast to the risk of aspiration or ejection of allergen in conventional i.n. challenge. Fifthly, using BMDCs instead of splenic DCs shows high productivity and good economic efficiency. From 1 mouse, sufficient BMDCs can be harvested to generate 100 asthma model mice; thus, BMDCs are more economical than splenic DCs. Sixth, mice need not to be anesthetized, which reduces stress in the animals and researchers. Finally, the administration of allergen-loaded materials (OVA-BMDCs) takes less time than conventional methods.
Interestingly, i.v. injection of OVA-BMDCs alone 3 times, without i.p. OVA sensitization, over 5 days elicited an asthma phenotype comparable to that induced by conventional 5 times i.n. administration of OVA over 26 days, although serum IgE and IgG were not elevated. Therefore, i.p. sensitization is required for antigen-antibody immune responses. In summary, i.p. OVA sensitization combined with i.v. OVA-BMDC transfer can be used to establish an ideal model of asthma in mice. The characteristics of the different protocols used to establish asthma murine models are described in Table.
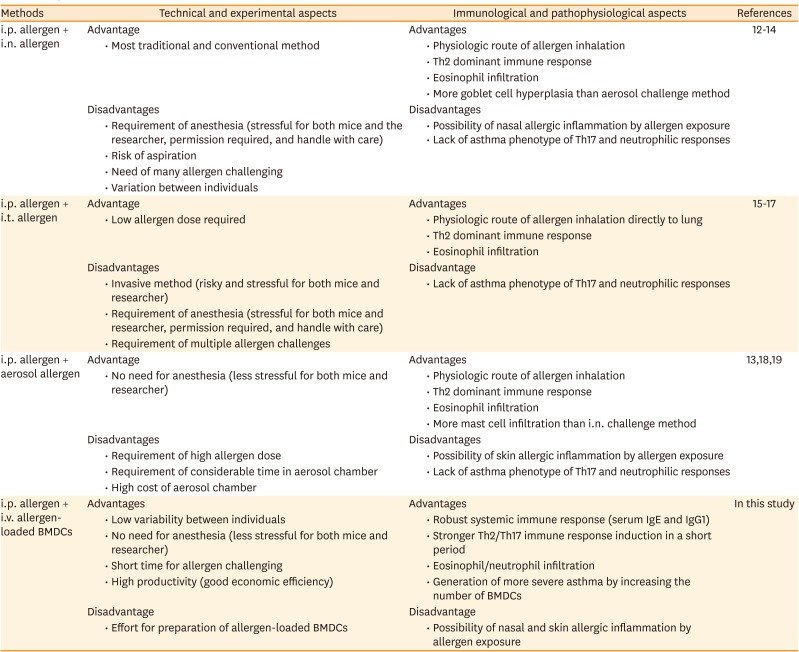
Table
Comparison of several methods to induce asthma in mice
| Methods | Technical and experimental aspects | Immunological and pathophysiological aspects | References | ||
|---|---|---|---|---|---|
| i.p. allergen + i.n. allergen | Advantage | Advantages | 121314 | ||
| • Most traditional and conventional method | • Physiologic route of allergen inhalation | ||||
| • Th2 dominant immune response | |||||
| • Eosinophil infiltration | |||||
| • More goblet cell hyperplasia than aerosol challenge method | |||||
| Disadvantages | Disadvantages | ||||
| • Requirement of anesthesia (stressful for both mice and the researcher, permission required, and handle with care) | • Possibility of nasal allergic inflammation by allergen exposure | ||||
| • Risk of aspiration | • Lack of asthma phenotype of Th17 and neutrophilic responses | ||||
| • Need of many allergen challenging | |||||
| • Variation between individuals | |||||
| i.p. allergen + i.t. allergen | Advantage | Advantages | 151617 | ||
| • Low allergen dose required | • Physiologic route of allergen inhalation directly to lung | ||||
| • Th2 dominant immune response | |||||
| • Eosinophil infiltration | |||||
| Disadvantages | Disadvantage | ||||
| • Invasive method (risky and stressful for both mice and researcher) | • Lack of asthma phenotype of Th17 and neutrophilic responses | ||||
| • Requirement of anesthesia (stressful for both mice and researcher, permission required, and handle with care) | |||||
| • Requirement of multiple allergen challenges | |||||
| i.p. allergen + aerosol allergen | Advantage | Advantages | 131819 | ||
| • No need for anesthesia (less stressful for both mice and researcher) | • Physiologic route of allergen inhalation | ||||
| • Th2 dominant immune response | |||||
| • Eosinophil infiltration | |||||
| • More mast cell infiltration than i.n. challenge method | |||||
| Disadvantages | Disadvantages | ||||
| • Requirement of high allergen dose | • Possibility of skin allergic inflammation by allergen exposure | ||||
| • Requirement of considerable time in aerosol chamber | • Lack of asthma phenotype of Th17 and neutrophilic responses | ||||
| • High cost of aerosol chamber | |||||
| i.p. allergen + i.v. allergen-loaded BMDCs | Advantages | Advantages | In this study | ||
| • Low variability between individuals | • Robust systemic immune response (serum IgE and IgG1) | ||||
| • No need for anesthesia (less stressful for both mice and researcher) | • Stronger Th2/Th17 immune response induction in a short period | ||||
| • Short time for allergen challenging | • Eosinophil/neutrophil infiltration | ||||
| • High productivity (good economic efficiency) | • Generation of more severe asthma by increasing the number of BMDCs | ||||
| Disadvantage | Disadvantage | ||||
| • Effort for preparation of allergen-loaded BMDCs | • Possibility of nasal and skin allergic inflammation by allergen exposure | ||||
i.p., intraperitoneal; i.n., intranasal; i.t., intratracheal; BMDCs, bone marrow-derived dendritic cells; Th, T-helper; Ig, immunoglobulin.
![]()
In the present study, we used the i.v. route (via the tail vein) to develop asthma, which can be inconsistent with the pathophysiology of asthma. However, the vast majority of mouse models rely on systemic sensitization of the immune system, although the site of allergen sensitization in humans is the airway itself.44 Moreover, Medoff et al.45 and Li et al.46 proved the feasibility of the i.v. route to induce allergic airway inflammation using T-cell transfer. According to Eggert et al.,47 i.v. injected BMDCs mainly accumulated in capillary-rich, well-perfused organs, such as the lungs and spleen, and MLNs. Komaru et al.48 applied i.v. administration of virally activated BMDCs to inhibit lung metastasis in a murine tumor model. The DCs administered via the tail vein migrated to the peritoneal vein and systemically circulated, and then entered the pulmonary vein and capillary and finally, the bronchial lumen by extravasation. The systemic immune response was directly driven by antigen-presenting DCs. The populations of inflammatory and goblet cells expanded in proportion to the number of OVA-BMDCs injected. Immunological parameters, including serum OVA-specific IgE, cytokines, and T-cell transcription factors supported good establishment of asthma. In this study, we aimed to develop a murine model of asthma with a stronger immune response using DCs. Furthermore, this DCs-based asthma model could be useful for immunological studies on the functions of DCs and T-cell responses in asthma. By controlling the characteristics and number of DCs, the T-cell immune response can be adjusted.
This study also has some limitations. First, an additional procedure for preparing allergen-loaded DCs is required. BMDCs were generated and cultured for 9 days; this in vitro procedure requires technical proficiency and is quite costly. Secondly, OVA is not physiologically relevant in humans. Nevertheless, this study provided proof of concept for i.v. transfer of OVA-BMDCs, and further studies are needed to investigate the asthma model using other allergens, such as house dust mite. Thirdly, our model shows enhanced systemic inflammation besides local airway inflammation. However, considering that this is a murine model that mimics human asthma and that mice do not develop human-like asthma naturally, systemic sensitization via an i.p. route is generally required to boost immune responses in order to generate asthma in mice. This could elicit systemic inflammation, although sensitization in humans is highly unlikely to occur through the peritoneum. This could be an inevitable characteristic and limitation of the currently available murine models. In the present study, although we have not evaluated GI tract or heart, other organs might also be affected by i.v. transfer of OVA-BMDCs. The enhanced systemic immune response is a special characteristic and could be an advantage of the present model because individuals with asthma sometimes exhibit features of systemic allergy with local allergy response. Moreover, systemic immune response could reflect local airway inflammation such as severity of asthma. For example, the serum level of allergen-specific IgE is a parameter of inflammation in asthmatic patients as well as asthmatic mice. Finally, this study does not clarify the detailed mechanism of Th2/Th17 induction by OVA-BMDCs in depth, since we focused mainly on generation of the model of asthma in this study. We are planning further studies to investigate the detailed mechanism of Th2/Th17 induction by OVA-BMDCs and the effect of other adjuvants in the asthma model.
Despite these limitations, the present study provided significant research advantages in that it established a reliable asthma model with low variation between individuals. Immune responses could be induced without anesthesia, and the degree of asthma inflammation can be controlled by varying the number of cells injected. Our model showed elevated Th17 immune responses as well as a Th2 response, including eosinophil and neutrophil infiltration, and was partially resistant to steroid administration. This is important because patients with severe or steroid-resistant asthma show mixed Th1/Th2/Th17 immune responses, and because neutrophil infiltration is related to a severe disease phenotype. Most of the murine asthma models developed to date show an allergic/eosinophilic phenotype, but lack a non-allergic/neutrophilic phenotype.21 Although a murine model of neutrophilic asthma was recently reported,49 it did not reflect heterogeneous asthma showing mixed eosinophilic/neutrophilic inflammation. Thus, the present model more accurately reflects the clinical features of heterogeneous human asthma and might offer an excellent experimental platform for the development of therapeutic strategies. Furthermore, this DCs-based asthma model can be used in further studies evaluating the effects of specific DC subsets or certain allergen-loaded DCs on asthma induction.
In conclusion, we developed a reliable and reproducible murine model of asthma for the first time using i.v. transfer of OVA-BMDCs. This model well mimicked heterogeneous human asthma. More severe asthma could be induced by increasing the number of cells transferred, and this model can be produced on a relatively large scale. Importantly, the good reproducibility, with low variation among individuals, and the fact that it does not require anesthesia support its usefulness for future studies on asthma. We expect that this i.v. OVA-BMDC-transfer murine model of asthma will contribute to the elucidation of the pathophysiology of this heterogeneous disease as well as to the identification of novel molecular targets for anti-inflammatory therapies for asthma.
 XML Download
XML Download