

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The most common type of kidney cancer is renal cell carcinoma, which arises from the epithelial cell lining of the proximal convoluted tubule [1]. Despite progress in systemic therapy of advanced renal cell carcinoma, the progression-free survival rate remains less than 1 year [2]. To overcome this poor prognosis, the therapeutic paradigm of advanced renal cell carcinoma has begun to shift in favor of precision medicine. Precision medicine is an approach to an individualized treatment model that seeks to identify effective therapy for every patient [3]. In this model, diagnostic testing is used to facilitate the selection of appropriate and optimal therapies based on an individual's genetic, molecular, or cellular signature. Owing to the intrinsic heterogeneity of renal cell carcinoma, precision medicine offers an attractive alternative to conventional therapies. Thus, development of a patient-derived in vitro model is essential for the realization of precision medicine in this diverse carcinoma.
Cancer cell lines and patient-derived xenografts are the most frequently used models in cancer research and anticancer drug screening for precision medicine [4]. Numerous studies have noted the limitations of two-dimensional (2D) culture of primary cell lines, especially in terms of their translation to human biology and treatment of tumors occurring in patients [56]. Cell-cell and cell-matrix interactions control cell phenotypes and function in vivo, making it essential to maintain these interactions in vitro to produce and retain cultured cell fidelity to normal phenotype and plasticity [7]. Patient-derived xenograft models address this concern but are plagued by their own limitations. These limitations include instability of engraft efficiencies and inaccurate reflections of the tumor microenvironment and immune system compared with those of the patient [8]. In addition, patient-derived xenograft models are costly and time-consuming, posing the most significant nonscientific barrier to the implementation of precision medicine for each carcinoma type [8]. Recently, cancer organoid cultures have been developed to establish in vitro growth of patient-derived samples at higher efficiency, and these systems are a promising alternative for use in fundamental cancer biology research as well as in large-scale drug screening [910].
Organoids are a miniature model of three-dimensional (3D) epithelial structure and recapitulate the histology and differentiation of the original organ [111213]. These systems are derived from a small number of cells from primary tissues, embryonic stem cells, or induced pluripotent stem cells. Importantly, organoids retain the functionality of their tissue of origin and are capable of self-renewal and selforganization [1415]. These characteristics of 3D organoid culture systems bridge the gap between traditional in vitro cell culture and in vivo patient-derived xenograft models [15]. In the present study, we establish a method for “patient-derived 3D organoid culture” from patients with advanced renal cell carcinoma. This study reveals the correlation between primary cancer tissue and patient-derived 3D organoid culture in this cancer type and provides the groundwork for future investigation of precision medicine for renal cell carcinoma.
MATERIALS AND METHODS
1. Human tissue
Normal renal tissues and renal cell carcinoma tissues were collected from patients who provided informed consent as stipulated by the Institutional Review Board (IRB) at Yonsei University (College of Medicine), and the study protocol was approved by the ethics committee (approval number: 4-2015-0104). Tissue samples were acquired from 13 patients who were enrolled in the study. The clinical data of the included patients are provided in Supplementary Table 1. Normal tissues were sampled at a distance from the tumor.
2. Organoid culture
For tissue dissociation, dissected human kidney tissue was minced with a blade to yield 1×1-mm pieces. Kidney tissues were incubated in 5 mL of Dulbecco's Modified Eagle's Medium (DMEM)/F12 (Gibco, Grand Island, NY, USA) supplemented with 1% fetal bovine serum (FBS), 3 mg/mL collagenase type II (Sigma-Aldrich, St. Louis, MO, USA), and 1× antibiotic antimycotic solution (Sigma-Aldrich) at 37℃ for 2 hours to digest tissue. After incubation, dissociated tissue was triturated vigorously by pipetting for 1 minute and then passed through a 70-µm cell strainer (Corning, New York, NY, USA). The cell pellet was centrifuged at ~200×g for 2 minutes and then gently washed with phosphate-buffered saline (PBS) twice. The cell pellets were resuspended in 1 to 2 mL of Matrigel (approximately 1×103 cells/µL of Matrigel; Corning) on ice and loaded in a 6-well plate. After polymerization of Matrigel, cells were incubated in YRC media, the organoid culture medium for kidney consisting serum-free keratinocyte medium (Gibco) supplemented with 0.01 mg/mL recombinant human insulin, 55 µg/mL human transferrin (substantially iron-free, Sigma-Aldrich), 5 ng/mL sodium selenite (ITS supplement, Sigma-Aldrich), 500 nM hydrocortisone (Sigma-Aldrich), 100 ng/mL human recombinant Noggin (PeproTech, Rocky Hill, NJ, USA), 10 nM leucine (Sigma-Aldrich), 5 µM Y-27632 (Enzo LifeSciences, Farmingdale, NY, USA), and 5% FBS. For serial passaging experiments, organoids were passaged at a 1:4 dilution every 1 to 2 weeks with 0.25% trypsin for 5 minutes at 37℃, followed by mechanical dissociation to nearly single cell suspensions. As a control group, a normal cell culture container was used, and the control group cells were serially passaged four times.
3. H&E staining
To characterize the 3D organoid culture and cell characteristics, a portion of the Matrigel under the 3D organoid culture was removed and re-solidified in 1.2% agarose in DMEM. After fixation in 10% formalin for 24 hours, samples were embedded in paraffin. Paraffin blocks were sectioned and adhered to slides. The slides were immersed in Xylene solution and then rehydrated by immersion in sequential ethanol dilutions. After soaking in hematoxylin solution for 8 minutes, slides were rinsed with running water. Finally, slides were incubated in eosin solution for 2 minutes and dehydrated using sequential ethanol and Xylene solution. DePeX (Serva, Heidelberg, Germany) solution was used for coverslip mounting, and slides were observed with an optical microscope.
4. Cell proliferation assay (CCK-8 assay)
Cells were loaded in triplicate in 48-well culture plates with Matrigel (12,000 cells/25 µL of each Matrigel dome). Cells were incubated for 72 hours in McCoy media and YRC organoid media, respectively. The cell proliferation was detected by use of cell counting kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan). To each well and plates 2-(2-Methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt was added and were incubated for 1 hour. A water-soluble formazan product in media was determined by using a Beckman Coulter microplate reader (Beckman Coulter, Miami, FL, USA) at 450 nm.
5. Immunofluorescence assay
The expression levels of carbonic anhydrase 9 (CA9), vimentin, and pan-keratin were compared by using immunofluorescence. Cancer cells and normal cells from patients were cultured in 8-well chamber slides (Labtech, East Sussex, UK). Each cell was seeded for 5 hours after trypsin treatment and removal of Matrigel. Cells were fixed with 4% paraformaldehyde solution for 10 minutes and then washed with PBS. Cells were then permeated in 0.5% Triton X-100 in PBS for 5 minutes, blocked in 5% bovine serum albumin (BSA) in PBS for 20 minutes, and incubated with diluted primary antibodies to CA9 (1:100; Novus Biologicals, Littleton, CO, USA), vimentin, and pan-keratin (Cell Signaling Technology, Danvers, MA, USA) in 5% BSA at 4℃ for 16 hours. Cells were washed with PBS and then incubated with secondary antibodies tagged with Alexa Fluor 488 or 594 (Molecular Probes, Eugene, OR, USA) at 25℃ for 1 hour (1:200). Images were acquired by laser scanning confocal microscopy (laser scanning microscope [LSM] Meta 700; Carl Zeiss, Oberkochen, Germany) and analyzed with LSM Image Browser software.
6. Immunohistochemical staining
Tissues from patients were embedded in paraffin, and 2-µm sections were prepared for immunohistochemical staining. After paraffin removal, the slides were pretreated in 0.01 M citric acid buffer (pH 6.0) for 10 minutes. After blocking with peroxidase blocking solution (Dako, Carpinteria, CA, USA), slides were incubated with primary anti-CA9, antivimentin, and anti-pan-keratin antibodies as described for immunofluorescence in 5% BSA (1:100) at 4℃ overnight. After washing in Tris-HCl buffer, the specific immunoreaction was detected using a secondary horseradish peroxidase-conjugated goat anti-rabbit antibody (1:200; Dako). After washing in Tris-HCl buffer, 3-3-diaminbenzidine tetrahydrochloride (DAB+; Dako) was used for visualization. Sections were counter stained with hematoxylin, dehydrated, mounted with DePeX (Serva), and examined with a microscope.
7. Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from organoids using TRIzol (Invitrogen [thermo fisher], Grand Island, NY, USA) and purified by use of RNeasy Mini Kits (Qiagen Korea, Seoul, Korea) in accordance with the manufacturer's protocol. For cDNA synthesis, 500 ng of RNA from each sample was reverse transcribed using the Superscript First Strand Synthesis system (Invitrogen). Quantitative real-time polymerase chain reaction (RT-PCR) was performed using SYBR green master mix reagent (Qiagen Korea) on an Applied Biosystem Step One Plus system (Applied Biosystems, Foster City, CA, USA). The cDNA samples were analyzed in triplicate for all analyses. To obtain relative values, the ΔΔCT method was used, and values were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression. Average values are shown. Primers were purchased from Qiagen Korea.
8. Western blot
Total protein extracts from organoids were prepared on ice using PRO-PREP protein extract solution (Intron, Seoul, Korea). Proteins from cell lysates were separated on a sodium dodecyl sulfate polyacrylamide gel and transferred to polyvinylidene fluoride membrane for 1 hour. Membranes were incubated at 4℃ overnight with anti-CA-9 and anti-GAPDH primary antibodies (Abcam, Cambrige, UK) diluted 1:1,000 with 5% BSA Tris-buffered saline-Tween 20 (TBS-T). After incubation, the membrane was washed with TBS-T, and then secondary antibodies (1:10,000; horseradish peroxidase-conjugated anti-mouse immunoglobulin G [IgG] or antirabbit IgG) were applied at room temperature for 1 hour. Labeled bands were detected by use of a West Pico chemiluminescent kit (Thermo Scientific, Rockford, IL, USA).
RESULTS
1. Morphologic characteristics of patient-derived 3D organoid culture of renal cell carcinoma
Hematoxylin and eosin (H&E)-stained samples of clear cell renal cell carcinoma showed the typical characteristics and morphology observed in the primary tissue (Fig. 1A). To evaluate whether 3D organoid culture retains the morphological characteristics of the primary tissue, organoid samples were examined under light microscopy. We observed that 3D organoid culture cells clustered to form a diverse colony that exhibited patterns similar to those found in the primary tissue (Fig. 1B). The samples were then embedded in paraffin and stained with H&E (Fig. 1C). Lipid-rich, clear cell morphology was readily observed, consistent with the primary tissue. In comparison to 2D culture, which shows morphologic denaturation after several passages (Supplementary Fig. 1), we observed that cell morphology in patient-derived 3D organoid culture was maintained (Supplementary Fig. 2). Thus, the main characteristics of primary clear cell renal cell carcinoma tissue were well maintained in organoid culture.
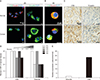
2. Validation of biomarkers of renal cell carcinoma in 2D culture, 3D organoid culture, and primary tissue
CA9, keratin, and vimentin were previously identified as biomarkers of renal cancer cells. To evaluate our 3D organoid culture system, we compared the expression patterns of these biomarkers between normal and cancer cells in 2D and 3D organoid culture as well as in primary tissue. Immunofluorescence analysis of 2D cultured renal cells revealed that CA9 and vimentin were more strongly expressed in cancer cells, whereas keratin was more strongly expressed in normal cells (Fig. 2A). The expression of CA9 and keratin was enhanced more in 3D organoid culture than in normal
cells (Fig. 2B). Consistent with our in vitro results, the expression of CA9 and vimentin in primary cancer tissue was stronger than in primary normal kidney tissue (Fig. 2C). Furthermore, the scores of CA9 and vimentin using immunohistochemical staining were highly positive in tissue samples from 5 patients with renal cell carcinoma (Fig. 2D). To quantify the increased expression of CA9 and vimentin in cancer cells, we performed RT-PCR analysis of normal and cancer tissues. The expression of both CA9 and vimentin was higher in cancer cells than in normal cells (Fig. 2E).
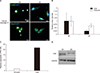
3. Increased expression of CA9 and vimentin in patient-derived 3D organoid culture of renal cell carcinoma
After establishing the validity of CA9, keratin, and vimentin as biomarkers in this model, we examined the differences in expression between 2D culture and 3D organoid culture. We found that the expression of CA9 and vimentin was significantly enhanced in 3D organoid culture compared with 2D culture. Immunofluorescence analysis showed that CA9 and vimentin were more strongly expressed in 3D organoid culture (passage 4) than in 2D culture (Fig. 3A). Additionally, the relative mean intensity score of CA9 and vimentin, as assessed by RT-PCR, was significantly higher in 3D organoid culture than in 2D culture (p<0.05; Fig. 3B). Furthermore, the fold upregulation in CA9 expression evaluated by RT-PCR indicated that the expression of CA9 was considerably increased in 3D organoid culture (Fig. 3C). The increased expression of CA9 in 3D organoid culture was confirmed by Western blot (Fig. 3D).
DISCUSSION
The advent of 3D organoid culture systems has advanced research on cancer, disease progression, metastasis, and treatment in many tissues, including the prostate, colorectal tissue, and liver. Prostate cancer is a leading cause of death among men in the United States and Western Europe [16]. A novel model of 3D organoid culture for advanced prostate cancer was reported in 2014 [17]. It was confirmed that this model reflects the nature of the original prostate cancer tissue. In addition to the models of organoid culture in colorectal cancer, models of hepatocellular carcinoma have been reported, which showed that the 3D organoid culture model preserves the properties of the original tissues well compared with the primary cancer tissue [1819]. Owing to the poor prognosis of patients with advanced renal cell carcinoma, development of a patient-specific precision model, such as 3D organoid culture, is imperative. However, there has been no previous report of development and validation of patient-derived 3D organoid culture.
In this study, we used our previous experience with a kidney tubular organoid model [20] to establish a technique for 3D organoid culture of advanced renal cell carcinoma to better represent the characteristics of the original patient-derived cancer cell. The gross and microscopic appearance of renal cell carcinoma is highly variable and poses a considerable challenge for culture models. With respect to the microscopic context, there are four major histologic subtypes of renal cell cancer: clear cell (conventional renal cell carcinoma, 75%), papillary (15%), chromophobe (5%), and collecting duct (2%) [21]. We focused on the clear cell subtype, as this type is the most commonly observed renal cell carcinoma. Our patient-derived 3D organoid culture was comparable to the histology observed in clear cell renal cell carcinoma (Fig. 1). We showed that the diverse clustering of 3D organoid culture exhibits patterns that resemble those of the original cell colony. Furthermore, we observed that the main characteristics of cell morphology, including a lipid-rich, clear cytoplasm and diverse colony formation, found in the primary tissue of clear cell renal cell carcinoma were well maintained in 3D organoid culture. Thus, patient-derived 3D organoid culture offers a potential biomimetic model of advanced renal cell carcinoma tissue.
CA9, keratin, and vimentin are well-known biomarkers in hypoxia and cancer metabolism, especially in renal cancer cells [2223]. CA9 and vimentin are known to be upregulated in cancer cells, whereas keratin expression is reduced. We had investigated our patient-derived 3D organoid culture system using these well-known cancer biomarkers. We found that both 2D culture and patient-derived 3D organoid culture of normal renal and renal cell carcinoma cells exhibited the expected patterns of expression of CA9, keratin, and vimentin. The increased expression of CA9 and vimentin and the concomitant decreased expression of keratin in cancer cells was maintained in our patient-derived organoid culture model. We further characterized the expression of these biomarkers in normal kidney tissue and renal cell carcinoma tissue and found that CA9 and vimentin were indeed expressed in cancer tissues as expected (Fig. 2C–E). Importantly, using immunofluorescence, RT-PCR, and Western blotting, we found that the expression of CA9 and vimentin was enhanced in patient-derived 3D organoid culture compared with 2D culture (Fig. 3). It is suggested that patient-derived 3D organoid culture is superior to 2D culture in terms of preserving the biomarker expression of the original cancer cells. Thus, our patient-derived 3D organoid culture model of clear cell renal cell carcinoma is consistent with patient biology, providing a robust model for additional study.
We note that there were several limitations to this study. The included number of patients in our study was small, and most of the samples were from low-stage tumors without any aggressive features, such as sarcomatoid change. We plan to continue to expand our study to include more samples from tumors with high-grade, aggressive phenotypes.
Despite the challenges to developing an organoid culture system for renal cell carcinoma, this patient-derived 3D organoid culture model may provide a new system for the development of a patient-specific in vitro clinical model of advanced renal cell carcinoma. The improved biological relevance of patient-derived 3D organoid culture makes it a promising model for fundamental cancer biology research as well as for large-scale drug screening. Future studies using this model may ultimately lead to the realization of precision medicine for patients with renal cell carcinoma. In this model, diagnostic testing is often used to select appropriate and optimal therapies in the context of a patient's genetic content or other molecular or cellular analysis.
CONCLUSIONS
We successfully established successful patient-derived organoid culture methods for advanced renal cell carcinoma. Our model retains the characteristics of advanced renal cell carcinoma with respect to morphology and biomarker expression. This patient-derived 3D organoid culture model may provide a new system for the development of patient-specific in vitro clinical models of advanced renal cell carcinoma.


 XML Download
XML Download