

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Lung cancer is the most commonly diagnosed cancer and the leading cause of cancer-related deaths worldwide.1 In addition, there is a 40% higher mortality rate in poorer areas of the United States compared to the most affluent counties.2 Although the underlying mechanisms of the initiation and development of lung cancer have been reported in the past decades, lung cancer prognosis and treatment still present significant challenges, with high relapse and drug-resistance rates reported among lung cancer patients.34 Therefore, further exploration of the mechanism and development of novel therapeutic methods for lung cancer treatment are required.
Over the past few decades, studies have mainly focused on the exploration of protein-coding genes, while noncoding RNA (ncRNA) was considered the result of transcription errors.5 However, growing evidence has revealed that ncRNAs can regulate cell proliferation, invasion, migration, and apoptosis and a remarkable variety of other biological functions related to tumors.67 MicroRNA (miRNA) is a class of short, single-stranded ncRNA that is <22 nucleotides in length. Recent study has shown that miRNA can modulate the expression of some key genes at both pre- and post-transcription levels. Additionally, the 3′ untranslated regions (3′UTR) of some messenger RNAs (mRNAs) and the process of translation can be modulated by miRNAs.5 In the past few years, miRNA regulation and function have attracted growing attention due to their involvement in many regulatory functions in cancer development. Recent study has revealed substantial aberrant expression of miRNAs in lung cancer tissues compared to adjunct normal tissues.8 In addition, several miRNAs have been reported to play vital regulatory roles in the onset and development of lung cancer. For instance, Li, et al.9 reported that miR-1254 can promote cell proliferation in lung cancer by binding to the 3′UTR of secreted frizzled related protein 1 (SFRP1) mRNA. The overexpression of stromal miR-143/145 can stimulate endothelial cell proliferation, which promotes neoangiogenesis in lung cancer development.10 By regulating the IL-6/STAT3 signaling pathway, miR-218 can inhibit lung cancer cell proliferation and invasion.11 The hsa-let-7g miRNA (also known as miR-let-7g and let-7g) is a member of the let-7 miRNA family that can exert vital biological effects in various diseases, including cancer. Hu, et al.12 reported that hsa-let-7g can exert anti-tumor effects in gastric cancer by inhibiting oncogenesis induced by oxidative stress. Another study indicated that hsa-let-7g could serve as an indicator for the prognosis of S-1-based chemotherapy.13 By binding to the 3′UTR of KRAS, hsa-let-7g can also promote the onset and progression of lung cancer in patients with chronic obstructive pulmonary disease.14 However, little is known about the role of hsa-let-7g in lung cancer cells.
The homeobox (HOX) genes encode a family of transcription factors that can bind to target genes sequence to modulate the expression level of target genes. HOXB1 is an important member of the HOX family, and several studies have demonstrated that HOXB1 is a vital regulator in the development of many diseases, including cancer. For example, the aberrant expression of HOXB1 contributes to the onset and development of glioma.15 HOXB1 is also an important factor in early vertebrate development, and HOXB1 dysregulation may result in congenital heart defects.16 Although numerous studies have reported crucial biological effects of HOXB1 in cancer development,1718 there has been no analysis of the role of HOXB1 in lung cancer.
In this study, we aimed to explore the correlation and mechanism between hsa-let-7g and lung cancer. Our analysis revealed significantly upregulated levels of hsa-let-7g in lung cancer tissues. The overexpression of hsa-let-7g may promote lung cancer development by directly decreasing the expression of HOXB1.
MATERIALS AND METHODS
Specimen collection and processing
The sample set included 20 cases of lung cancer and 20 adjacent normal lung tissues, which were obtained from surgical operation and verified by postoperative pathology. The removed tissue was washed using DPEC water. Target tissues were cut using special scissors, transferred to refrigerated tubes, and stored in a liquid nitrogen tank. Using the WHO criteria for the classification of digestive tumors, all of the sampled patients were staged. Before collecting the samples, we obtained consent from the Ethics Committee of The Second Clinical Medical College of Yangtze University (IRB No.: 2018111029).
Cell culture
Both A549 and H1944 lung cancer cells were purchased from the Shanghai cell bank. Cells were maintained in RPMI-1640 medium, which contained 10% fetal bovine serum (Wisent, Quebec, Canada) and 100 u/mL streptomycin/penicillin at 37℃. Cells were cultivated in a humidified incubator containing 5% carbon dioxide.
Plasmid construction and transfection
An inhibitor of hsa-let-7g (LV-inhibitor) and a negative control lentivirus (LV-NC) were designed as short hairpin RNA (shRNA) sequences, and were cloned by and purchased from Genepharm (Hangzhou, China). After adding 1 µg/mL polybrene (Genepharm) to each well (1×105 cells/well), cells were incubated for 0.5 h at 37℃ before the addition of 15 µL of transfection regent and lentivirus vector. The transfected cells were selected with puromycin (2.0 µg/mL) for 14 days. Fluorescence was detected after 48 hours. Five colonies were selected, and numbered as colonies 1 to 5. After checking the effect of shRNA transfection by qRT-PCR, colony 2 was selected for downstream experiments.
RNA extraction and qRT-PCR
Total mRNA was extracted from A549 and H1944 lung cancer cells and tissues using TRIZOL Reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer's instructions. We evaluated the amount and quality of extracted RNA using a Nanodrop 2000 spectrophotometer. Using a Reverse Transcription Kit (Takara, Dalian, China), the extracted RNA was transcribed into complementary DNA. Subsequently, qRT-PCR was performed to measure the level of mRNA in tissues and cells through SYBR II Premix Taq (Takara), and all processes were completed according to the instructions of Prime Script RT-PCR Kit (Takara Biochemicals, Tokyo, Japan). GAPDH was used as the internal reference. The primers utilized are listed in Table 1.
Western blot
Total protein from A549 and H1944 lung cancer cells was extracted using radioimmunoprecipitation assay buffer (RIPA, Beyotime Biotechnology, Shanghai, China) and quantified by bicinchoninic acid (BCA) protein assay kit (KeyGen BioTech, Nanjing, China), according to the manufacturer's instructions. Subsequently, electrophoresis was performed, and the protein was transferred to polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA) at 300 mA for 1.2 h. The membranes were immersed in 5% non-fat milk for 1 hour, and then incubated with primary antibodies against HOXB1 (1:1000, Santa Cruz, Dallas, TX, USA) or GAPDH (1:1000, Santa Cruz) at 4℃ overnight. After washing the membranes three times with TBS-T, appropriate secondary antibodies were added and the membranes were incubated for 1 hour at 20℃. Then, a chemiluminescence detection kit (Thermo Fisher, Waltham, MA, USA) was used to develop the blots.
Cell viability assay
Cells were transferred into a 96-well plate and cultured with a cell counting kit-8 (CCK-8) reagent (Dojindo, Kumamoto, Japan). Briefly, 10 µL CCK-8 was added to each well at 24, 48, 72, 96, and 120 hours. A microplate reader (Tecan, Mechelen, Belgium) was then used to detect the absorbance value (OD) of each well at 450 nm. Additionally, the proliferation of A549 and H1944 lung cancer cells was detected by EdU assay. Each experiment was independently performed three times, and the mean values are presented.
Colony formation assay
The transfected A549 and H1944 cells were seeded into 6-well plates at a density of 1000 cells per well, and then cultured for 7 days. After formation of visible colonies, 2 mL of 4% paraformaldehyde was added to each well and incubated for 15 min. After washing with PBS, the wells were stained with a Giemsa stain kit for 30 min. The colonies were then counted using an ordinary optical microscope.
Apoptosis detection
Cultured cells were collected into a cell suspension through trypsinization, and the density was adjusted to 5×105 cells/mL after washing twice with cold PBS. Then, 10 µL of Annexin V-FITC was added to 100-µL cell suspension in each tube, and cells were incubated in the dark for 15 min at 4℃. Flow cytometry analysis was performed immediately after the addition of 380-µL binding buffer and 10-µL propidium iodide.
Luciferase activity assay
The wild-type (WT) or mutant (MUT) hsa-let-7g sequences encoding the binding site of HOXB1 were designed and subcloned into pGL3 basic vector (Promega, Madison, WI, USA). A549 cells were seeded on 24-well plates for 24 hours. Mimics of hsa-let-7g were co-transfected with 10-µg pLUC-WT-HOXB1 or pLUC-MUT-HOXB1 using Lipofectamine 2000 reagent (Invitrogen). Luciferase activity was measured using Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
SPSS 19.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 5.0 software (GraphPad Software Inc., San Diego, CA, USA) were used to perform statistical analysis. Data are presented as the mean±standard deviation. Student t-test analysis was performed to compare quantitative variables. Statistically significance was defined as p<0.05.
RESULTS
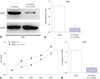
Overexpression of hsa-let-7g was detected in lung cancer tissues
To investigate the expression profile of hsa-let-7g in lung cancer tissues and adjacent normal tissues, we extracted mRNA from the 20 pairs of tissues samples and detected the expression level of hsa-let-7g by qRT-PCR. We observed significantly increased hsa-let-7g expression in lung cancer tissues compared to the adjacent normal tissues (Fig. 1A). The pathological features of the 20 lung cancer patients were then compared to hsa-let-7g expression level, and we found significantly increased expression level of hsa-let-7g in patients with higher tumor classification (Fig. 1B). Additionally, the expression level of HOXB1 was measured, and significantly downregulated HOXB1 levels were observed in cancer tissues (Fig. 1C). All of these results were statistically significant.
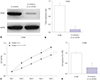
Silencing of hsa-let-7g expression resulted in decreased proliferation and increased apoptosis in both A549 and H1944 cells
To further investigate the regulatory mechanism of hsa-let-7g in lung cancer cells, a lentivirus-mediated inhibitor of hsa-let-7g (LV-inhibitor) was tested in A549 and H1944 lung cancer cells. Transfection efficiency was measured by qRT-PCR, and significantly decreased hsa-let-7g expression level was observed after transfection with the hsa-let-7g LV-inhibitor (Fig. 2A and E). CCK-8 assay and colony formation assay showed suppression of cell proliferation in the hsa-let-7g LV-inhibitor group in both A549 and H1944 cells compared to non-transfected cells (Fig. 2B, C, F, and G). Moreover, cell apoptosis was significantly promoted in the hsa-let-7g LV-inhibitor group (Fig. 2D and H). These results suggested that hsa-let-7g promotes lung cancer progression.
hsa-let-7g targets HOXB1
A recent study reported that has-let-7g can promote the development of osteosarcoma by targeting HOXB1.19 We predicted a similar mechanism to exist in lung cancer. We used the TargetScan website (http://www.targetscan.org) and confirmed HOXB1 as a direct target of hsa-let-7g, with the target binding site in the 987-994 nt region of 3′ UTR in HOXB1 mRNA. The binding was re-confirmed by reporter assay using luciferase (Fig. 3A and B). We further analyzed the correlation between HOXB1 and hsa-let-7g by measuring the expression level of HOXB1 mRNA by qRT-PCR, and measuring the protein expression level by Western Blot in A549 and H1944 cells transfected with the hsa-let-7g LV-inhibitor or a negative control. The results revealed increased expression levels of HOXB1 mRNA and protein after silencing hsa-let-7g (Fig. 3C–F).
hsa-let-7g regulates A549 and H1944 cells viability and apoptosis by regulating HOXB1
To further investigate whether hsa-let-7g regulates the development of lung cancer by targeting HOXB1, we next performed a co-transfection assay. The increased HOXB1 protein expression level induced by transfection with the hsa-let-7g LV-inhibitor was significantly reversed by co-transfection of hsa-let-7g LV-inhibitor and an inhibitor targeting HOXB1, LV-sh-HOXB1, in both A549 and H1944 cells (Fig. 4A, 4B, 5A, and 5B). This suggested that hsa-let-7g modulates HOXB1 expression level in lung cancer cells. Additionally, CCK-8 and apoptosis assays showed increased cell viability and decreased apoptosis after co-transfection, which differed from cells that were transfected with only the hsa-let-7g LV-inhibitor (Fig. 4C, 4D, 5C, and 5D). All of these results suggested that hsa-let-7g may promote proliferation and inhibit apoptosis in lung cancer by modulation of HOXB1 expression level.
DISCUSSION
In this study, we investigated a potential role of hsa-let-7g in lung cancer. We found significantly increased levels of hsa-let-7g in lung cancer tissues and downregulated expression of HOXB1. The overexpression of hsa-let-7g may promote lung cancer development by directly decreasing the expression level of HOXB1. HOXB1 is a vital regulator in cancer development,1718 and this is the first study to suggest a role of HOXB1 in lung cancer.
Previous studies have shown key roles of miRNAs in both physiological and pathological processes.2021 In particular, miRNAs can modulate the initiation and development of lung cancer. Xu, et al.22 reported that increased expression of miR-4326 was positively associated with lung cancer cell proliferation. Analysis of clinical data of 154 non-small-cell lung cancer patients and 63 benign lung disease patients by Jiang, et al.23 revealed that miR-26b can act as a diagnostic and prognostic biomarker for lung cancer. Another miRNA, miR-1284, can inhibit cell proliferation, induce apoptosis, and exert anti-tumor effects in lung cancer.24 In this study, we observed that hsa-let-7g miRNA exerted oncogenic influence in A549 and H1944 lung cancer cells.
The hsa-let-7g is a vital member of the miRNA let-7 family, and several studies have reported vital roles of hsa-let-7g in the onset, progression, and migration of various cancers. For instance, has-let-7g can decrease the expression level of oncogene Myc, increase tumor suppressor gene p16 expression, and inhibit hepatocellular carcinoma cells proliferation.25 Additionally, by targeting HOXB1, hsa-let-7g can activate the NF-κB pathway that contributes to the initiation of osteosarcoma.19 Also, hsa-let-7g plays vital roles in breast cancer migration by negatively regulating FOXC2.26
Previous studies motivated us to explore the role of hsa-let-7g in lung cancer cells, and we predicted that miRNA could be a potential therapeutic target. In our study, we used qRT-PCR to measure the expression profile of has-let-7g in 20 paired samples, and found a higher expression level in lung cancer tissues compared to that in normal tissues. In addition, we found a positive association between tumor classification and hsa-let-7g expression level by analyzing pathological data of the patients. An inhibitor of hsa-let-7g was used to detect biological function in A549 and H1944 lung cancer cells. CCK-8 and apoptosis assays revealed that silencing hsa-let-7g decreased cell viability and induced apoptosis, suggesting that the overexpression of hsa-let-7g in lung cancer patients may similarly inhibit apoptosis and promote tumor initiation. Acting as a sponge of mRNA is the primary mechanism by which miRNAs exert regulatory effects,27 and TargetScan analysis identified potential binding sites between has-let-7g miRNA and HOXB1 3′ UTR, and luciferase activity assay confirmed the direct targeting of HOXB1 by has-let-7g. Zhou, et al.19 previously pointed out that has-let-7g could exert oncogenic effects in osteosarcoma by binding the 3′ UTR of HOXB1. We predicted that hsa-let-7g may regulate the initiation and progression of lung cancer by targeting HOXB1. The inhibition of has-let-7g resulted in increased expression levels of HOXB1 mRNA and proteins. Co-transfection of has-let-7g LV-inhibitor and LV-sh-HOXB1 confirmed that has-let-7g promoted lung cancer development by targeting HOXB1, since decreasing HOXB1 could reverse the effect.
HOXB1 is an anti-tumor gene that belongs to the HOX family, and is located in chromosome 17q21.32. Growing evidence has indicated that the deregulation of HOXB1 makes vital contributions to the progression of various diseases, including cancers.2829 HOXB1 can exert tumor suppressive effects by inhibiting the expression of survival oncogenic genes.30 Han, et al.15 reported that low expression level of HOXB1 contributed to cell invasion and proliferation, and inhibited apoptosis in glioma.
In this study, upregulated HOXB1 was observed in A549 and H1944 lung cancer cells after transfection with hsa-let-7g LV-inhibitor. Additionally, the knockdown of HOXB1 contributed to cell viability by inhibiting apoptosis, which was the opposite of the effect observed when hsa-let-7g was inhibited.




 XML Download
XML Download