

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Oxidative stress is related to an imbalance between the production of reactive oxygen species (ROS) and free radicals as well as inappropriate antioxidant function. The ROS, formed by an inadequate one-electron defect of oxygen, play critical roles in homeostasis and cell signaling. Hydrogen peroxide (H2O2), a highly reactive ROS, has been commonly used to mimic in vitro oxidative stress in many types of cells [1,2].
Bone is an essential structure to support the body and has its own homeostatic activity to maintain its integrity. This activity has been called a bone remodeling process regulated by the balanced action of osteoblasts and osteoclasts [34]. Osteoblasts, which produce and release matrix proteins, have a critical role in the process of bone formation. Intracellular accumulation of ROS in osteoblasts leads to the destruction of bone homeostasis [5].
Dexmedetomidine, a specific agonist of the α2-adrenoceptor, has been commonly used in operating rooms and intensive care units for analgesia and sedation [67]. Recently, dexmedetomidine has been reported to have a protective effect on the lung, intestine, liver, and kidney against the oxidative stress that is triggered by endotoxemia or ischemia reperfusion [891011].
Thus, we aimed to investigate whether dexmedetomidine has a protective effect against H2O2-induced oxidative stress and to elucidate the mechanism of H2O2-induced cell death in normal human fetal osteoblast (hFOB) cells.
MATERIALS AND METHODS
1. Reagents
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT) was purchased from Sigma (St. Louis, MO, USA). Antibodies against Collagen type I (Col I), bone morphogenetic protein-2 (BMP-2), osterix, and transforming growth factor-β (TGF-β) were purchased from Abcam (Cambridge, MA, USA). GAPDH, mouse anti-rabbit IgG antibody, and rabbit anti-mouse IgG antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All other chemicals and reagents were purchased from Sigma unless otherwise specified.
2. Cell culture
Human fetal osteoblast cell line (hFOB 1.19) was purchased from ATCC (Rockville, MD, USA). Cells were cultured in Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F-12) with 4 mM L-glutamine, 1.5 g/L sodium bicarbonate, 4.5 g/L glucose, and 1.0 mM sodium pyruvate supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin (GIBCO-BRL, Rockville, MD, USA). Cells were incubated at 37℃ in a humidified 5% CO2–95% air incubator.
3. Treatment of dexmedetomidine
The stock of dexmedetomidine (490 µM) was kept frozen at -20℃ until use and diluted to the designated concentration with DMEM/F-12 when needed. Prior to dexmedetomidine treatment, cells were grown to about 75% confluence and then exposed to dexmedetomidine at various concentrations (0, 0.1, 1, 5, and 10 µM) for 2 h to determine the most appropriate concentration. The cells were divided into the following three groups: control group—the cells were incubated at 37℃ in a humidified atmosphere with 5% CO2 without dexmedetomidine treatment, H2O2 group—the cells were exposed to 200 µM of H2O2 for 2 h, and Dex/H2O2 group—the cells were pretreated with 5 µM of dexmedetomidine for 2 h and then exposed to 200 µM of H2O2 for 2 h.
4. Cell viability assay
The viability of hFOB cells was determined using the MTT assay. Cells were cultured in 96-well plates (4 × 103 cells/well). The cells were then treated with the indicated concentrations of drug. At the endpoint of treatment, 100 µl of MTT (500 mg/ml) was added to each well. The cells were incubated for 4 h at 37℃. The formazan crystals, which were formed, were then solubilized in DMSO (200 µl/well) by constant shaking for 15 min. The cell viability was measured by an ELISA reader (Tecan, Männedorf, Switzerland) at 620 nm excitatory emission wavelength.
5. Fluorescence microscopy
Cells were harvested and cytocentrifuged onto a clean, glass slide. Cells were stained in 1 µg/ml Hoechst 33342 for 15 min at 37℃ in the dark and washed in phosphate buffered saline (PBS). The slides were mounted with glycerol. The samples were observed and photographed under an epifluorescence microscope (Carl Zeiss, Goettingen, Germany).
6. Flow cytometer analysis
Quantification of apoptotic cells was determined by Annexin V-FITC/PI staining. The adherent cells were collected by centrifugation and then resuspended in 500 µl of 1X binding buffer. Then, they were stained with 5 µl Annexin V-FITC and propidium iodide (PI) (50 µg/ml) and incubated at room temperature for 5 min in the dark. The cells were analyzed with a CYTOMICS FC 500 flow cytometer system (Beckman Coulter, CA, USA ). The results were shown as quadrant dot plots with intact cells (Annexin V-/PI-), early apoptotic cells (Annexin V+/PI-), late apoptotic cells (Annexin V+/PI+), and necrotic cells (Annexin V-/PI+).
7. Alizarin red S staining
Mineralization of hFOB cells was determined in 24-well plates using Alizarin red S (Sigma, St. Louis, MO, USA) staining. The cells were washed once with distilled water after fixation with 4% paraformaldehyde and then stained with 2% alizarin red solution according to the manufacturer's protocol.
8. Western blot analysis
The cells (1.5 × 106) were washed twice in ice-cold PBS, resuspended in 200 µl ice-cold solubilizing buffer (300 mM NaCl, 50 mM Tris-Cl (pH 7.6), 0.5 % Triton X-100, 2 mM PMSF, 2 µl/ml aprotinin, and 2 µl/ml leupeptin), and incubated at 4℃ for 1 h. The lysates were centrifuged at 13,200 rpm for 30 min at 4℃. Protein concentrations of cell lysates were determined using a Bradford protein assay (Bio-Rad, Richmond, CA, USA), and 20 µg of protein was resolved by a 10% SDS/PAGE gel. The gel was transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). After blocking, the membranes were reacted with appropriate primary antibodies. Immunostaining with secondary antibodies was detected using SuperSignal West Femto substrate (Pierce, Rockford, IL, USA).
RESULTS
1. Dexmedetomidine inhibits H2O2-induced cell death in hFOB cells
The effect of dexmedetomidine on hFOB cells was demonstrated over various concentrations (0, 0.1, 1, 5, and 10 µM). The viability of hFOB cells pretreated with dexmedetomidine was not changed compared to the control. Dexmedetomidine did not show any cytotoxic effect on the hFOB cells (Fig. 1A). In contrast, after treatment with a wide range of H2O2 concentrations (0, 25, 50, 100, 200, and 400 µM) for 2 h, the cell viability was significantly decreased in a dose-dependent manner (Fig. 1B). All subsequent experiments were performed with 5 µM of dexmedetomidine and 200 µM of H2O2. To compare the cell viability among the three groups, we performed another MTT assay. We pretreated with 5 µM of dexmedetomidine for 2 h and then exposed hFOB cells to 200 µM H2O2 for 2 h. As shown in Fig. 2, cell viability was markedly decreased in the H2O2 group compared to the control, while viability was significantly improved by dexmedetomidine pretreatment. As a result, we discovered that dexmedetomidine pretreatment significantly protected the hFOB cells from H2O2-induced cell death.
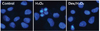
2. Dexmedetomidine protected hFOB cells against H2O2-induced cell death.
The mechanism of H2O2-induced cell death and protective effect of dexmedetomidine on hFOB cell were investigated by Hoechst 33342 and Annexin-V FITC/PI staining. Cells were observed under a fluorescence microscope (× 400) (Fig. 3). In the control and Dex/H2O2 groups, nuclei were lightly stained by the dye and morphological changes were not detected. In contrast, the nuclei of H2O2 treated cells presented typical apoptotic features, which are fragmented and deeply stained nuclei. We next investigated whether H2O2-induced cell death through an apoptotic mechanism. Annexin-V FITC/PI staining was used for the detection of apoptotic stages. Compared with the control group, in the H2O2 group the portion of Annexin-V(+)/PI(+) cells increased from 1.2% to 25%. However, pretreatment with dexmedetomidine 2 h prior to H2O2 treatment significantly attenuated the percentage of Annexin-V(+)/PI(+) cells to 10.0 % (P < 0.05), demonstrating the anti-apoptotic effect of dexmedetomidine (Fig. 4).
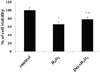
3. Positive effects of dexmedetomidine on mineralization of hFOB cells.
Mineralization of hFOB cells was investigated using Alizarin red S staining. This staining clearly showed that dexmedetomidine pretreatment enhanced the staining intensity of hFOB cells when compared to that of the H2O2 group (Fig. 5).
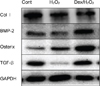
4. Effect of dexmedetomidine on expression of bone-related protein in hFOB cells.
We examined the activation of bone-related protein in hFOB cells by western blotting analysis. The recruitment of osterix to the membrane occurs via a Col I-dependent mechanism; thus, Col I is essential for bone formation in vivo. The levels of Col I, BMP-2, osterix, and TGF-β were increased in the Dex/H2O2 group compared to the H2O2 group (Fig. 6).
DISCUSSION
Various external stimuli can trigger oxidative stress by toxins, ischemia, or mechanical stress. Oxidative stress has previously been regarded as an inducer of a variety of bone diseases including osteoblastic apoptosis and osteoporosis [1213]. However, the biological mechanism through which oxidative stress affects osteoblast death is still not fully understood.
The goals of this study were to determine the protective effect of dexmedetomidine on hFOB cells that have undergone H2O2-induced oxidative stress and to investigate the regulation of bone homeostasis. Our findings showed that dexmedetomidine treatment did not cause any cytotoxic effects in hFOB cells (Fig. 1A). We also utilized the cell viability assay to demonstrate the toxicity of H2O2. As a result, H2O2 treatment induced cell death in a dose-dependent manner. In particular, a concentration of more than 100 µM caused oxidative stress in hFOB cells (Fig. 1B). To investigate the effect of dexmedetomidine against H2O2-induced cell death, we made a comparison among the three distributed groups: control, H2O2, and Dex/H2O2. The viability of the H2O2 treatment group was decreased compared to that of the control. However, pretreatment with dexmedetomidine restored the cell viability that had been decreased by H2O2.
Cell staining with Hoechst confirmed that pretreatment with dexmedetomidine reduced H2O2-induced apoptosis (Fig. 3). Hoechst staining can distinguish dead cells from live cells by indicating fragmentation and condensation of chromatin, which causes the cells to brightly fluorescence [14]. We explored this process in more detail using Annexin V-FITC/PI double staining. One event in the earlier stage of apoptosis includes translocation of phosphatidylserine (PS) from the inside of the cell membrane to the surface. Annexin V has high affinity for PS, and fluorochrome-labeled Annexin V can be used for the detection of PS translocation using flow cytometry. PI is frequently used as a nuclear staining dye. The ability of PI to enter a cell is dependent on the membrane permeability; PI cannot get into live or early apoptotic cells because of the intact cell membrane. In late-stage apoptosis and necrotic cells, the intact part of plasma and nuclear membranes deteriorates, allowing PI to enter the membranes and display fluorescence [151617181920]. In the H2O2 group, the portion of Annexin-V(+)/PI(+) cells was increased. This suggested that H2O2 affects the late stage of apoptosis and dexmedetomidine has a protective effect against H2O2-induced apoptosis in hFOB cells (Fig. 4).
Osteoblasts, derived from mesenchymal stem cells, are responsible for the formation of bone extracellular matrix and are able to mineralize into bone matrix [212223]. Alizarin red S staining suggested that the bone mineralization matrix was weak in H2O2 groups. As expected, dexmedetomidine pretreatment enhanced the mineralization staining (Fig. 5). Next, we examined the recovery effect of dexmedetomidine on the expression levels of osteoblastic differentiation markers (Col I, BMP-2, osterix, and TGF-β) in hFOB cells by western blot analysis. Expression of all osteoblastic markers was decreased in the H2O2 group while dexmedetomidine enhanced their expression (Fig. 6).
In conclusion, our results suggest that H2O2-induced cell death via apoptotic signaling in hFOB cells and pretreatment with dexmedetomidine can prevent H2O2-induced oxidative stress. In addition, dexmedetomidine enhanced the activity of osteoblastic differentiation in response to oxidative injury. We propose the potential therapeutic value of dexmedetomidine in treating oxidative stress via inhibiting apoptosis and enhancing osteoblast activity. Moreover, the current investigation could be further evidence to support the antioxidant potential of dexmedetomidine in vitro.



 XML Download
XML Download