

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Among women, endometrial cancer is the sixth most common cancer worldwide [1], increasing its importance particularly in the developed countries reflecting their lifestyle changes. Traditionally, endometrial cancer has been classified into types 1 and 2 based on etiology and clinical behavior [23]. Type 1 tumors are estrogen-dependent well-differentiated adenocarcinomas, arising in the background of endometrial hyperplasia, comprising up to 80% of endometrial cancer cases. Contrarily, type 2 tumors are estrogen-independent, poorly differentiated adenocarcinomas, which occur de novo from atrophic endometria. Over many years, histological types, lymph node (LN) and distant metastasis have been regarded as the critical prognosis factors of endometrial cancer [45]. Since 2013, the classification of endometrial cancer has been drastically changed [6]; the importance of the tumor's genomic features in determining its aggressiveness and resistance to therapy has become apparent. Thus, to develop new therapeutic strategies, further investigation of tumor-driving genes is the pressing issue today.
Laminins are heterotrimeric extracellular matrix proteins that are composed of the alpha, beta, and gamma chains, which mediate cellular signaling through interaction with cell membrane receptors [7]. The effect of specific laminin isoform expressions on cancer progression has been vigorously studied; for instance, the LAMC2 gene, encoding the laminin subunit gamma 2 chain, is an established prognostic marker of various types of cancer [8-10]. In this study, we focused on the LAMC1 gene encoding the laminin subunit gamma 1 (LAMC1) chain, which has recently been reported to be involved in the carcinogenesis of fallopian tube cancer [11]. LAMC1 overexpression has been reported to be correlated with poor prognosis of hepatocellular carcinoma [1213] and meningioma [14], but its role in endometrial cancer remains to be elucidated.
By investigating the LAMC1 expression patterns in clinical specimens, we show that overexpression of LAMC1 in endometrial cancer is related to aggressive histological types, LN metastasis, advanced International Federation of Gynecology and Obstetrics (FIGO) stage, and poor prognosis. We also examine the downstream molecular changes caused by the overexpression of this new potential therapeutic target.
MATERIALS AND METHODS
1. Patients and tissue samples
Endometrial tissues were obtained from 97 patients with endometrial carcinoma and from 9 patients with atypical endometrial hyperplasia who underwent surgery at Keio University Hospital (Tokyo, Japan) from 1999 to 2013. All specimens were fixed in 10% phosphate-buffered formalin and embedded in paraffin. The 4 μm-thick sections were stained with hematoxylin and eosin to confirm the presence of tumor and to assess its histological characteristics. Written informed consent was obtained from each participant regarding the use of samples for research. The Ethics Committee of Keio University approved this study (approval No. 20130336).
2. Immunohistochemistry
The specimens were deparaffinized in xylene and rehydrated in a graded series of ethanol. Antigen retrieval was performed with citrate buffer (pH 6.0) for 20 minutes at 95°C. After blocking of endogenous peroxidase activity with 0.3% H2O2 in phosphate-buffered saline, the specimens were incubated overnight with anti-LAMC1 antibody (HPA001908, 1:500; Sigma-Aldrich, St. Louis, MO, USA) at 4°C. Indirect immunohistochemical staining was performed using N-Histofine Simple Stain MAX-PO (Nichirei Biosciences Inc., Tokyo, Japan). The specimens were counterstained with hematoxylin, dehydrated in a graded series of ethanol, dried and coverslipped.
3. Evaluation of immunohistochemical staining
Although laminin gamma 1 is a member of the extracellular matrix, the LAMC1 antibody stains its precursor, which localizes to the cytosol. We defined overexpression of LAMC1 as >10% of tumor cells or normal endometrial cells showing higher immunoreactivity compared to their stromal counterparts in 5 high-power fields in each section. Two investigators (H.K. and Y. K.) independently evaluated the slides in a blinded manner.
4. Cell line and culture
We used 2 human endometrial cancer cell lines: HEC50B cells were purchased from the American Type Culture Collection (Manassas, VA, USA); SPAC-S cells were kindly provided by Dr. Tokuichi Kawaguchi (Japanese Foundation for Cancer Research, Cancer Institute, Tokyo, Japan). HEC50B cells were maintained in Dulbecco's modified eagle medium supplemented with 10% fetal bovine serum (FBS). SPAC-S cells were cultured in 20% MCDB152 medium and 80% DM160 medium supplemented with 5% FBS, insulin-transferrin-selenium (5 μg/mL; Collaborative Research Co., Lexington, MA, USA), and epidermal growth factor (10 ng/mL; Takara Biomedicals, Shiga, Japan) [15].
5. siRNA LAMC1 inhibition
We purchased siRNA duplexes from Thermo Fisher Scientific; Silencer® Select siLAMC1 (siRNA ID s8077) and SilencerTM Select Negative Control No.1 siRNA (product No. 4390843) (Thermo Fisher Scientific, Waltham, MA, USA). Twenty-four hours after seeding, cells were transfected with siRNA duplexes for 48 hours using the Lipofectamine RNAiMAX reagent (Invitrogen, Waltham, MA, USA).
6. Microarray analysis
Total RNA from the HEC50B and SPAC-S cells was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany) in combination with RNase-free DNase (Qiagen). The quality of RNA was assessed using the Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA). The RNA samples were labeled using Low Input Quick Amp Labeling Kit (one-color; Agilent) and then hybridized to the Agilent SurePrint G3 Human Gene Expression version 3.0 Microarray (Agilent) according to the manufacturer's instruction. The arrays were scanned using the Agilent DNA microarray scanner G2505C (Agilent). The statistical software R (version 3.4.1; R Foundation, Vienna, Austria) and the Bioconductor package limma were used to process the raw data. Briefly, the raw data underwent background correction by the “normexp” method, followed by quantile normalization across samples. Probes that displayed low expression levels across all samples were excluded from the analysis. The empirical Bayes method implemented in limma [16] was used to identify genes that were differentially expressed (false discovery rate [FDR] <0.01) between the control (control siRNA) and the experimental (LAMC1 siRNA) groups. We performed gene ontology (GO) analysis using the Database for Annotation, Visualization and Integrated Discovery version 6.8 (DAVID, http://david.abcc.ncifcrf.gov). The p<0.05 was considered statistically significant.
Furthermore, we performed hierarchical clustering using R and identified differentially expressed genes (DEGs) possibly involved in cancer progression by referring to GeneCards (https://www.genecards.org) and The Human Protein Atlas (https://www.proteinatlas.org).
7. Quantitative real-time polymerase chain reaction (RT-PCR) analysis
Total RNA was obtained from cells as mentioned earlier, and 500 ng of each sample was reverse-transcribed using SuperScript VILO Master Mix (Thermo Fisher Scientific). RT-PCR analysis was performed in triplicate using SYBR Green Master Mix and Thermal Cycler Dice (Takara Biomedicals). The primer sets for the RT-PCR are listed in Supplementary Table 1.
8. Statistical analysis
Statistical analysis was performed using SPSS statistical software version 24.0 (IBM Japan, Tokyo, Japan). Categorical variables were reported as absolute numbers and percentages. Continuous variables were reported as median and range or as mean and standard deviation. Correlations between overexpression of LAMC1 and clinicopathological factors were evaluated by χ2 test. We calculated the progression-free and overall survival from the date of surgery to either the last follow-up or date of recurrence or death. The Kaplan-Meier method was used to estimate the probability of progression-free and overall survival, and log-rank test was used to compare between groups. The p<0.05 was considered statistically significant.
RESULTS
1. LAMC1 is overexpressed in endometrial carcinoma
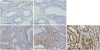
The representative LAMC1 immunohistochemical staining results are shown in Fig. 1. In normal proliferative phase endometrium (obtained from the unaffected area of the resected specimen) and atypical endometrial hyperplasia, we observe only slight LAMC1 staining in the endometrial stroma and vascular endothelia, and LAMC1 expression in the endometrium did not exceed that in the stroma. On the contrary, the tumor cells in endometrial carcinoma (endometrioid carcinoma grades 1 and 3, and clear cell carcinoma) showed prominent LAMC1 expression, clearly exceeding that in the stroma. As described in Table 1, none of the atypical endometrial hyperplasia specimens showed LAMC1 overexpression (0/9), whereas overexpression of LAMC1 was observed in 76.3% (74/97) of the endometrial carcinoma specimens, representing a significant difference (p<0.001).
Fig. 1
LAMC1 expression pattern in endometrial tissues. (A) Normal endometrium (proliferative phase). (B) Atypical endometrial hyperplasia. (C) Endometrioid carcinoma grade 1. (D) Endometrioid carcinoma grade 3. (E) Clear cell carcinoma. (A,B) There were evaluated as negative. (C-E) There were evaluated as positive for LAMC1 overexpression. Scale bars, 100 μm.
LAMC1, laminin subunit gamma 1.

2. Relationship between LAMC1 overexpression and clinicopathological features in endometrial carcinoma
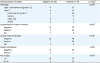
We analyzed the relationship between LAMC1 overexpression and clinicopathological features in patients with endometrial cancer. The age at diagnosis and body mass index did not differ among patients with or without LAMC1 overexpression (Supplementary Table 2). Focusing on the histological type, type 2 cases had a significantly higher LAMC1 overexpression rate (88.7% [47/53], p=0.002) than type 1 cases (61.4% [27/44]). Although grade 3 endometrioid carcinoma (81.5% [22/27]) had a higher LAMC1 overexpression rate than grades 1–2 (61.4% [27/44]), no statistical significance was noted among these 2 groups (p=0.113). Of note, in serous carcinoma and clear cell carcinoma, which are especially aggressive histological types, nearly all of the cases had LAMC1 overexpression (95.0% [17/18] and 100% [5/5], respectively). Furthermore, lymphovascular space invasion (LVSI), LN metastasis, and advanced FIGO stage were related to a significantly higher LAMC1 overexpression rate, while distant metastasis was not (Table 2).
Table 2
Relationship between overexpression of LAMC1 and clinicopathological factors
3. Association between LAMC1 overexpression and patient prognosis in endometrial carcinoma
The follow-up period of the 97 patients ranged from 3 to 122 months, with a median period of 81 months. A total of 22 patients experienced relapse of the disease, and 19 died. Patients with endometrial carcinoma overexpressing LAMC1 tended to have a shorter progression-free survival (p=0.201), and strikingly, had a significantly poor overall survival (Fig. 2, p=0.045).
Fig. 2
Kaplan-Meier curves representing the association between LAMC1 overexpression and (A) PFS and (B) OS rates. Log-rank tests indicated that patients with overexpression of LAMC1 had a poorer overall prognosis.
LAMC1, laminin subunit gamma 1; OS, overall survival; PFS, progression-free survival.

4. LAMC1 silencing in endometrial cancer cell lines
Several previous studies have indicated that LAMC1 silencing results in impaired proliferation, migration, and invasion in various cancer cell lines [131718]. However, the molecular mechanisms underlying these changes remain to be investigated. Thus, we aimed to identify genes influenced downstream of LAMC1 overexpression by manipulating its expression in endometrial cancer cells.
LAMC1 overexpression was prominent among type 2 endometrial carcinomas. Therefore, we chose HEC50B, which represents poorly differentiated endometrioid carcinoma [19], and SPAC-S, which represents serous carcinoma [15], to evaluate the effects of LAMC1 inhibition. The cells were transfected with either control siRNAs or siRNA targeting LAMC1, and total RNA was provided to microarray gene expression assays.
The results of the microarray analysis revealed that a total of 652 genes were differentially expressed (fold change >2, FDR <0.01) commonly in HEC50B and SPAC-S in response to LAMC1 knockdown, including 211 upregulated and 441 downregulated genes. Supplementary Fig. 1 shows the results of the GO term enrichment analysis. As to the biological process, the downregulated DEGs significantly enriched in cell cycle, posttranscriptional regulation of gene expression, and translation, suggesting that LAMC1 silencing may negatively impact endometrial cancer progression through these changes. Supplementary Fig. 2 presents the top 100 influenced genes showing common expression changes in HEC50B and SPAC-S. Of note, 8 independently expressed genes that are correlated with tumor progression according to GeneCards and the Human Protein Atlas [20] were influenced commonly in HEC50B and SPAC-S. Four favorable prognostic factors, LZTFL1, TAPT1, SEL1L, and PAQR6, were upregulated, whereas 4 unfavorable prognostic factors, NME7, TMEM109, CCDC58, and ANKRD40, were downregulated by LAMC1 knockdown (Table 3), suggesting their role in the tumor-promoting effect of LAMC1 overexpression. Quantitative PCR confirmed the expression changes of the aforementioned 8 genes by LAMC1 knockdown (Fig. 3).
Table 3
Genes correlated with tumor progression that are influenced commonly in HEC50B and SPAC-S by LAMC1 knockdown

Fig. 3
Relative expression of genes in HEC50B and SPAC-S transfected with siRNA targeting LAMC1. Cells were transfected with either LAMC1 or control siRNAs for 48 hours. Silencing of LAMC1 and expression changes of genes identified by microarray analysis were confirmed. The p-values are as shown (Student's t-test). All quantitative data are expressed as means± standard deviation (n=3 experiments).
LAMC1, laminin subunit gamma 1.

DISCUSSION
In our present study, while none of the atypical endometrial hyperplasia specimens had LAMC1 overexpression, endometrial carcinoma had a significantly high LAMC1 overexpression rate, suggesting the involvement of LAMC1 in the carcinogenesis of endometrial cancer. Because LAMC1 was reported to be overexpressed in the serous tubal intraepithelial carcinoma [11], we first expected it to be an early diagnostic marker for endometrial cancer. However, given the fact that type 1 endometrial carcinoma has a relatively low LAMC1 overexpression rate and that type 2 endometrial carcinoma possesses a higher LAMC1 overexpression rate than type 1, LAMC1 may not be the initial driver gene of oncogenesis in endometrial carcinoma. Instead, it seems to be correlated with its aggressiveness. In line with this idea, our analysis indicated that endometrial carcinoma overexpressing LAMC1 is likely to have LVSI, LN metastasis, and advanced FIGO stage. Together with the fact that LAMC1 knockdown suppressed cell motility and invasion in endometrial cancer cells [17], our results suggest a relationship between LAMC1 overexpression and lymphatic invasion.
LAMC1 overexpression was correlated with a significantly poor overall survival in endometrial carcinoma. In addition, progression-free survival showed the same tendency but without statistical significance, and this discrepancy may be based on the low aggressiveness of LAMC1-negative endometrial carcinoma, allowing the cure or remission of the recurrent disease with subsequent therapy.
The downstream mechanism through which LAMC1 influences tumor aggressiveness is largely unknown. Here, we reported that knockdown of LAMC1 in endometrial cancer cells caused significant expression changes in LZTFL1, TAPT1, SEL1L, PAQR6, NME7, TMEM109, CCDC58, and ANKRD40 at the transcription levels. LZTFL1 is a tumor suppressor gene identified through the analysis of the hotspot of tumor suppressor genes at chromosome 3p21.3, and the lack of its expression is correlated with poorly differentiated histological types, metastasis, and poor prognosis in patients with gastric cancer [21]. In vitro studies have revealed that overexpression of LZTFL1 causes suppression of epithelial-mesenchymal transition, cell migration and invasion in cancer cells. Although the majority of its functions remain unclear, the interaction of LZTFL1 and beta-catenin nuclear translocation has been reported recently [22]. SEL1L is the human homolog of the Caenorhabditis elegans sel-1 gene, which acts as a tumor suppressor in breast cancer and pancreatic ductal adenocarcinoma. Overexpression of SEL1L inhibits tumorigenicity in vitro and in vivo through the modulation of the Notch receptor genes, Smad4, activin A and PTEN [23-26]. NME7 is a member of the NME/NDK family, which mainly acts as nucleoside-diphosphate kinases and protein histidine kinases. NME7 is widely known for its microtubule-organizing functions and role in ciliary cargo transport [27], and its participation in cancer biology has been overlooked. Its role in the chemosensitivity of myeloma was reported only recently [28]. The interaction between LAMC1 and these genes, and the specific molecules involved in the exhibition of tumor aggressiveness in LAMC1-overexpressing tumors, is to be further investigated.
In summary, our data indicate that overexpression of LAMC1 is strongly associated with histological type, LVSI, LN metastasis, advanced FIGO stage, and poor overall survival in endometrial cancer. These results suggest that LAMC1 is a potent biomarker for identifying patients needing aggressive adjuvant therapy. Gene expression microarray analysis elucidated 8 candidate genes (LZTFL1, TAPT1, SEL1L, PAQR6, NME7, TMEM109, CCDC58, and ANKRD40) that may be involved in cancer progression in the downstream of LAMC1 expression. Further elucidation of the underlying mechanism should lead to the discovery of new therapeutic targets.
 XML Download
XML Download