

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Although the breast cancer death rate has decreased rapidly from 1989 to 2015, breast cancer is still the second most common leading cause of cancer mortality among women in the United States [12]. Currently, surgery, radiotherapy, chemotherapy, and hormone therapy are the main therapeutic approaches used to treat breast cancer, with targeted therapy and immunotherapy as co-treatment options; however, their efficiencies are limited in the case of breast cancer recurrence and metastasis progression [3]. Therefore, it is necessary to discover novel molecular targets and their related signaling pathways to suppress carcinogenesis and tumor metastasis.
The N-myc and STAT interactor (NMI) is an interferon-γ inducible gene product that interacts with several key molecules in cancer cell signaling such as N-myc, C-Myc, SOX10, TIP60, and all STATs except STAT2 [45678]. The role of NMI in tumorigenesis, cancer progression, and metastasis remains unclear. A recent study showed that high expression of NMI predicts poor prognosis and promotes tumor growth in glioblastoma [9]. Moreover, NMI is overexpressed in metastatic hepatocellular carcinoma and promotes hepatocellular carcinoma progression via the BDKRB2 and MAPK/ERK pathways [10]. By contrast, some reports suggest that NMI is induced in response to cellular stress, and a fraction of NMI is translocated into the nucleus to stabilize ARF (INK4a/ARF), a tumor suppressor, and aid in the stabilization of TP53 [11]. Subsequent signaling studies also revealed that NMI expression negatively regulates oncogenic Wnt/β-catenin signaling [12]. Some studies showed that loss of NMI had negative impacts on STAT5-driven expression of the transforming growth factor β (TGFβ) signaling repressor SMAD7, which induced epithelial-mesenchymal transition (EMT). Silencing NMI expression in epithelial-like breast cancer cell lines induces molecular markers and morphological attributes of the mesenchymal-like phenotype and promotes the invasive ability of these cells [13]. As advanced invasive breast cancer progresses, NMI expression is reduced, and thus the absence of NMI may indicate poor prognosis in breast cancer [131415]. Moreover, NMI was reported to have a vital role in modulating drug response in cells through the activation of autophagy in breast cancer cells [16].
In this study, we investigated NMI protein expression in breast cancer using a tissue microarray (TMA) and estimated the clinicopathologic significance of NMI as a prognostic factor. We also evaluated the association between expression and prognostic significance of NMI according to the molecular subgroup of breast cancers. Furthermore, we validated NMI expression significance in another independent cohort, The Cancer Genome Atlas (TCGA) database. Our data demonstrate that NMI expression is a potential prognostic factor for breast cancer patients and suggests an important role of NMI in aggressive tumor behavior and the progression of breast cancer.
METHODS
Patient selection and study design
A total of 382 consecutive breast cancer patients who underwent curative-intent resection in 2008 at Seoul National University Hospital, Seoul, Korea, were enrolled in the study. Primary treatments included radical mastectomy, modified radical mastectomy, breast-conserving surgery, and sentinel lymph node biopsy or axillary lymph node dissection. All cases were invasive carcinoma of no special type according to the World Health Organization classification [17]. Patients who had received neoadjuvant chemotherapy were excluded. Formalin-fixed, paraffin-embedded tissues of 382 patients with breast cancer were used to construct the TMA. Clinicopathologic parameters and patient survival data were reviewed and collected via an electronic medical records system. The anatomic tumor stage was determined according to the 8th criteria of the American Joint Committee on Cancer [18]. Histologic grading was performed according to the Nottingham grading system [19]. The requirement for informed consent from the patients was waived by the ethics committee. This study was approved by the Institutional Review Board (IRB) of the Seoul National University Hospital (IRB No. 1512-076-728).
Immunohistochemistry
Immunohistochemical staining was performed for the molecular classification of breast cancers on whole tissue sections according to the guidelines introduced in the 13th St. Gallen International Breast Cancer Conference [20]. The immunohistochemical staining for the estrogen receptor (ER; 1:100, 1D5; Novocastra Laboratories, Newcastle, UK) and progesterone receptor (PR; 1:200, PgR636; Dako, Glostrup, Denmark) expression was counted and categorized as positive when ≥ 1% of the tumor cell nuclei were stained, according to the 2010 American Society of Clinical Oncology/College of American Pathologists (ASCO/CAP) guidelines [21]. The immunohistochemical expression of the human epidermal growth factor receptor 2 (HER2; 4B5; Ventana, Tucson, USA) was assessed based on the 2013 ASCO/CAP guidelines. The presence of HER2 amplification was detected with an additional fluorescence in situ hybridization assay using the PathVysion HER2 DNA Probe Kit (Abbott Molecular, Downers Grove, USA) [22]. The Ki67 (1:100, MIB-1; Dako) proliferative index was also assessed. The immunohistochemical staining for NMI (1:75, Novus biologicals, Centennial, USA) expression was evaluated based on the percentage and intensity of cytoplasm-stained tumor cells. NMI expression was considered high when ≥ 40% of tumor cells were stained. All immunohistochemically stained slides were reviewed by 2 experienced breast pathologists (MSJ and HSR) for improved accuracy.
In vitro cell line culture
We performed western blot analyses of NMI expression in 13 different breast cancer cell lines and 1 normal breast epithelial cell line (MCF10A). Some of these cell lines (MCF7, T47D, BT474) were ER-positive, others (SkBr3, MDA-MB-453) were HER2-positive, and the rest (BT-20, MDA-MB-468, HCC38, MDA-MB-157, MDA-MB-436, MDA-MB-231, Hs578T, BT-549) were triple-negative breast cancer (TNBC) cell lines. Cell lines with triple-negative status are differentiated as basal A and basal B cell lines, with basal A being more luminal-like and basal B being more basal-like [23]. All cell lines were grown in Dulbecco's Modified Eagle Medium supplemented with 10% fetal bovine serum (GenDEPOT, Katy, USA) and 1% penicillin-streptomycin (Gibco, Grand Island, USA). All cell lines were cultured at 37°C in a humidified atmosphere under 5% CO2.
Western blot analysis
For intracellular protein extraction, the transfected cells within a monolayer were lysed in ice-cold T-PER buffer (Thermo Fisher Scientific, Waltham, USA) containing a protease inhibitor cocktail (Roche, Basel, Switzerland). The isolated proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and were transferred to a Polyvinylidene fluoride membrane (Millipore, Billerica, USA). After blocking with 5% skimmed milk in Tris-buffered Saline with Tween 20 (TBST), the membrane was incubated with primary antibodies specific to NMI (1:1,000, Novus biologicals) and β-actin control (1:1,000, C-2; Santa Cruz Biotechnology, Santa Cruz, USA). After incubation with primary antibodies, the membrane was washed in TBST for 5 minutes and then 3 times for 10 minutes each. The membrane was incubated with secondary antibodies (horseradish peroxidase-conjugated goat anti-rat immunoglobulin G (H + L), Thermo Fisher Scientific) for 1 hour at room temperature (20–22°C). The membrane was washed 3 times in TBST for 10 minutes each.
The TCGA survival data analysis
The NMI expression data of breast cancer samples (n = 1,075) from the TCGA cohort were evaluated using the Human Protein Atlas database (the HPA program, http://www.proteinatlas.org/), a biomarker discovery strategy using antibody-based proteomics. The NMI expression levels were divided into 2 groups (high expression and low expression). The Kaplan-Meier (log-rank) test for p-value was used to show the resulting analysis of the correlation between gene expression and patient survival.
Statistical analysis
The χ2 and Fisher's exact tests were performed to evaluate the association between clinicopathologic categorical variables and protein expression. The student's t-test was used to compare the continuous variables. Kaplan-Meier survival curves with log-rank tests were used to analyze the time to progression and death. Multivariate analysis was performed using the Cox proportional-hazards model. Statistical analyses were performed using the IBM SPSS v22.0 (IBM Corp., Armonk, USA). A p < 0.05 was considered statistically significant.
RESULTS
Clinicopathologic characteristics
In total, 382 cases of breast cancer patients were enrolled. Most of the clinicopathologic parameters are listed in Table 1. The patients were aged between 24 and 78 years old (mean 48.6 ± 10.2 years). The tumor size ranged from 0.4 cm to 12.2 cm (mean 2.46 ± 1.30 cm). We classified 264 patients (69.1%) as “luminal A”, 17 patients (4.5%) as “luminal B”, 34 patients (8.9%) as “HER2-positive”, and 67 patients (17.5%) as “triple-negative” based on the results of the immunohistochemical staining. The median follow-up time was 72.8 months (3.5–83.8 months). During the follow-up, 49 patients (12.8%) had recurrence or metastasis, and 3 patients (0.8%) died.
Table 1
Correlation between clinicopathologic characteristics and NMI expression with breast cancer
Correlation between NMI expression and clinicopathological parameters
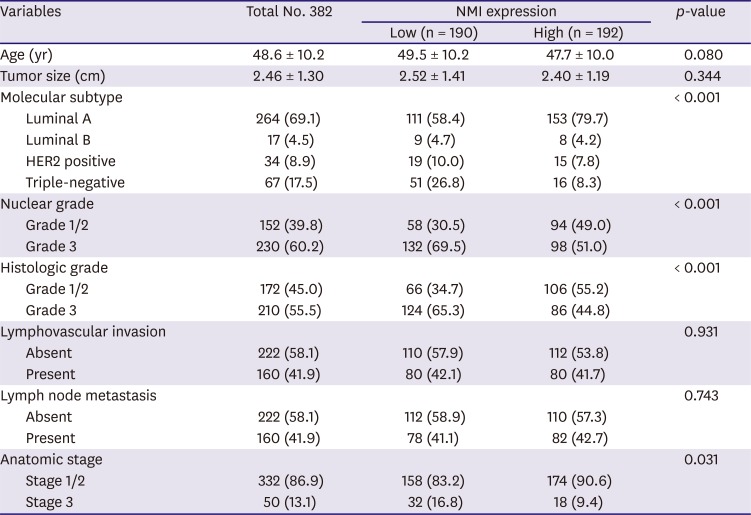
NMI protein expression was observed in normal breast epithelial cells, mainly in the cytoplasm (Figure 1). In the breast cancer samples, the tumor cells either showed cytoplasmic NMI expression or a loss of expression (Figure 1). High NMI expression was observed in 192 cases (50.3%) and low NMI expression was observed in 190 cases (49.7%). Low NMI expression was frequently observed in the “triple-negative” molecular subtype and showed statistical significance (p < 0.001). Low expression of NMI was also significantly associated with conditions of high nuclear grade (p < 0.001), high histologic grade (p < 0.001), and advanced anatomic stage (p = 0.041), is shown in Table 1.
Figure 1
Representative immunohistochemical results for NMI expression in normal breast tissue and breast cancer. (A) NMI expression is observed in the cytoplasm of normal breast epithelial cells (IHC for NMI, 400× magnification). (B, C) Loss of NMI expression and presence of NMI expression in breast cancers (IHC for NMI, 4× and 400× (inlet) magnification, respectively).
NMI = N-myc and STAT interactor; IHC = immunohistochemistry.
Prognostic significance of NMI expression
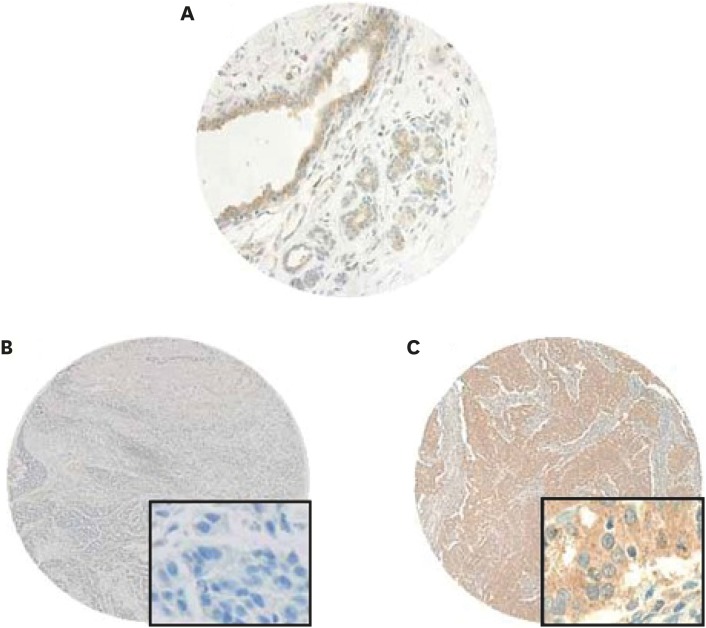
The Kaplan-Meier survival analysis showed that low NMI expression was significantly associated with poor progression-free survival (PFS; p = 0.038) in breast cancer (Figure 2). We further analyzed patient prognosis and NMI expression in the molecular subgroups of breast cancer and found that low NMI expression was not significantly associated with patient prognosis in all the molecular subgroups of breast cancers (Figure 3).
Figure 2
Kaplan-Meier survival analyses for the NMI expression as a determinant of PFS.
NMI = N-myc and STAT interactor; PFS = progression-free survival.
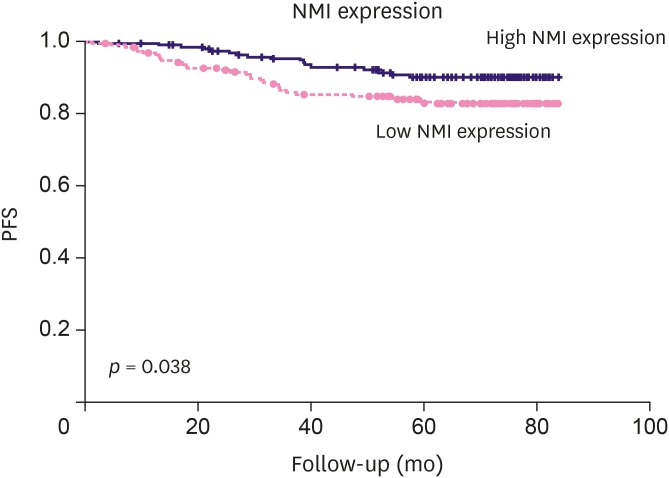
Figure 3
NMI expression and patient PFS analysis according to each molecular subtype. (A) Luminal A; (B) Lumina B; (C) HER2 positive; (D) Triple-negative breast cancer.
NMI = N-myc and STAT interactor; PFS = progression-free survival; HER2 = human epidermal growth factor 2.
Univariate Cox regression analyses revealed that young patient age (hazard ratio [HR], 0.386; p = 0.003), larger tumor size (HR, 4.320; p < 0.001), high nuclear grade (HR, 3.793; p = 0.001), high histologic grade (HR, 4.734; p < 0.001), presence of lymphovascular invasion (HR, 2.956; p = 0.001), presence of lymph node metastasis (HR, 1.868; p = 0.030), high anatomic stage (HR, 3.997; p < 0.001), and low NMI expression (HR, 0.546; p = 0.041) were independent poor prognostic factors for PFS (Table 2).
Table 2
Univariate and multivariate Cox regression analyses for PFS
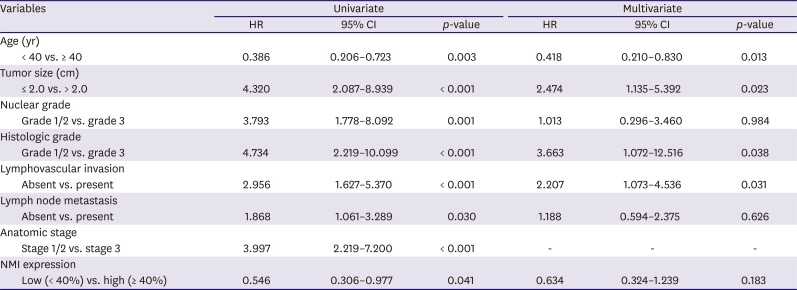
In the multivariate analysis, young patient age (HR, 0.418; p = 0.013), larger tumor size (HR, 2.474; p = 0.023), high histologic grade (HR, 3.663; p = 0.038), and presence of lymphovascular invasion (HR, 2.207; p = 0.031) were significant factors associated with PFS (Table 2).
NMI expression in breast cancer cell lines
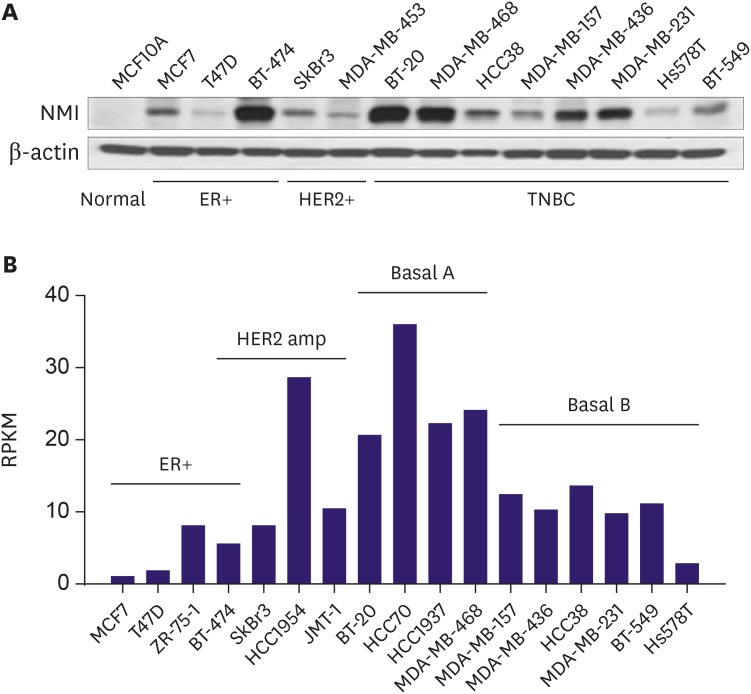
We analyzed the NMI protein levels in 13 different breast cancer cell lines and a normal breast epithelial cell line. As shown in Figure 4A, some of these cell lines (MCF7, T47D, SkBr3, MDA-MB-453, HCC38, MDA-MB-157, Hs578T, BT-549) showed a noticeable reduction in NMI expression. However, there were some cell lines (BT-474, BT-20, MDA-MB-468, MDA-MB-436, MDA-MB-231) that retained NMI expression. Upon a closer look at the breast cancer subgroups according to the molecular subtypes, the ER-positive and basal B type of TNBC subgroups showed a significant reduction in NMI expression (Figure 4).
Figure 4
(A) Western blot analysis shows that NMI expression is reduced in some of the breast cancer cell lines. (B) Subgroup analyses of NMI expression levels in breast cancer cell lines.
NMI = N-myc and STAT interactor; HER2 = human epidermal growth factor 2; ER = estrogen receptor; RPKM = reads per kilobase of transcript per million mapped reads; TNBC = triple-negative breast cancer.
Validation of NMI expression and its prognostic significance
The HPA program revealed that from 1,075 breast cancer cases, 829 cases (77.1%) showed high NMI expression and 246 cases (22.9%) showed low NMI expression. The median follow-up time was 2.37 year. The analysis showed that patients with low NMI expression levels tended to be associated with lower survival probability compared with those with high NMI expression (p = 0.053).
DISCUSSION
It has been demonstrated that NMI is widely expressed in fetal and adult tissues, and overexpressed in multiple cell lines; however, its functional role in cancer is still controversial [4]. Recently, some studies demonstrated that high expression of NMI is associated with tumor progression and predicts poor prognosis [910]. By contrast, it has been reported that NMI inhibits tumor growth by upregulating DKK1 in breast cancer cell lines [12]. Li et al. [24] demonstrated that NMI is a vital component of a transcriptional factor tricomplex that contains breast cancer type 1 and c-Myc, which may function as a tumor suppressor by regulating c-Myc-induced hTERT promoter activity in breast and ovarian cancers. It is also reported that NMI expression is negatively correlated with the stage and grade of breast tumors [13]. However, there is little information on their predictive value in the prognosis of breast cancer patients. In the present study, we found that low NMI expression was significantly associated with high nuclear and histologic grade, and anatomic stage. Low NMI expression was also significantly associated with poor PFS. Therefore, we suggest that low NMI expression is a potential prognostic factor in breast cancer patients. Additionally, we observed that low NMI expression levels tended to associate with lower survival probability in the TCGA cohort using the HPA program.
Some reports have suggested that loss of NMI promotes EMT by the activation of TGFβ/SMAD signaling in breast cancer [1315]. EMT is a unique process of epithelial cells gradually changing into a mesenchymal cell phenotype, resulting in enhanced motility and invasiveness [2526]. Recent studies indicated that EMT is relevant to the invasion and metastasis of TNBC [2728]. Because of the lack of targeting agents and limited therapeutic options, treatment of TNBC remains an important clinical challenge [29]. In this study, low NMI expression was more frequently observed in the “triple-negative” molecular subtype. Using in vitro analyses, the reduction of NMI expression was frequently found in the ER-positive and basal B type of TNBC subgroups. The basal B type cell lines, designated the mesenchymal cluster or normal-like/claudin-low, overexpress genes associated with invasive and aggressive tumor features, and cancer stemness [23]. Samant and Shevde [30] demonstrated that NMI silencing had noticeable effects on cellular morphology that indicated the acquisition of a mesenchymal-like phenotype. Devine et al. [13] also reported that cell lines that predominantly displayed epithelial characteristics retain expression of NMI whereas mesenchymal-like cell lines display a noticeable loss of NMI protein expression. These findings suggested that NMI expression was not only associated with molecular subtype (hormone receptor and/or HER2 expression status) but also with cellular morphology. In our analysis, ER-positive breast cancer cell lines, excluding the basal B type, also showed low NMI expression. The TCGA data suggested that luminal A, luminal B, and normal-like breast cancer subtypes (ER+/PR+ status) showed lower NMI messenger RNA levels. Further studies should be performed to understand the association of molecular subtype, cellular morphology, and NMI expression.
When we further correlated NMI expression with patient prognosis in the “triple-negative” subgroup, low NMI expression was not significantly associated with patient PFS (p = 0.825). However, in this study, the number of TNBC cases was limited. Further studies are required to determine the prognostic significance of low NMI expression in the molecular subtypes of breast cancer, especially TNBC.
In conclusion, we suggest that low NMI expression is associated with poor prognosis of breast cancer patients. Additionally, we found that low NMI expression is frequently observed in TNBC, especially in more mesenchymal-like and aggressive cell lines. Therefore, NMI expression could be a useful prognostic biomarker and a potential novel therapeutic target in breast cancer.
 XML Download
XML Download