

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Influenza virus is an infectious virus that infects 5%–15% of the global population every year, causing serious respiratory illnesses and resulting in about half million of deaths [1]. Vaccination is the most effective way to prevent the disease by influenza virus infection. Since influenza viruses continue to evolve by antigenic drift as RNA viruses, World Health Organization updates influenza vaccine components twice a year to match antigenicity with the circulating viruses [2]. Recently, influenza vaccines are mainly quadrivalent vaccines containing two A type (H1N1 and H3N2) and two B type (B/Yamagata and B/Victoria lineage) viruses and most of them are manufactured by growing influenza viruses using eggs in a traditional manner. However, propagating the influenza virus in the egg can produce virus variants that are different from those cultured in animal cells, resulting in changed antigenicity from the original virus derived from the patient [345]. In particular, candidate vaccine virus (CVV) for vaccine production is mainly produced through passaged culture in eggs, where the antigenicity change may occur due to egg adaption of the virus. This adaptation is the result of several amino acid mutations in hemagglutinin protein. Especially, since the amino acid in which the mutation occurs is a receptor binding site of the hemagglutination (HA) protein or a position very close thereto, the amino acid sequence change at this position may lead to a change in antigenicity [67].
Since the human case was confirmed in 1968, H3N2 virus has been more genetically and antigenically evolved than other types of virus so that the vaccine component was updated more than 30 times [8910]. In particular, low vaccine effectiveness were identified for H3N2 virus in the last 2012–2013 season, which was reported to be due to the mutation of HA protein by egg adaptations in CVV used in vaccine manufacturing rather than to the antigenic drift of virus [11]. At that time, the CVV for A/Victoria/361/2011 (H3N2) was subjected to amino acid mutations in three position of HA protein by egg adaptations, which were all areas corresponding to the antigenic site as H156Q, G186V, and S219Y. All of these amino acid mutations had increased effects on α2,3-linked receptors and decreased on α2,6-linked receptors in terms of binding avidity. Especially in the case of G186V, it is known to be a mutation that is characteristically found in the egg adaption of the influenza virus [1213]. The low vaccine effectiveness of H3N2 was also reported in the 2016–2017 season, which is also due to the change of glycosylation site depending on amino acid mutation of HA antigen by egg adaptation [1415]. In the case of clade 3C.2a virus recommended as a vaccine strain, a glycosylation site was added according to the K160T amino acid mutation. But egg-adapted vaccine strain of 3C.2a lacked this glycosylation site, and thus the vaccine effectiveness was lowered even though it was matched well with the actual circulation strain [16]. The correlation between glycosylation and antigenicity of HA protein is well known and influenza virus can avoid host immune system by adding N-linked glycosylation site to HA [17]. H3N2 viruses have evolved more actively through mutations in the antigenic sites of HA proteins, and these amino acid mutations result in the addition of new glycan, which significantly affects antibody binding. Over the last 50 years, more than 10 new N-glycosylation sites have been added to the H3N2 viral HA antigen, and the addition of these sites has particularly affected the activity of receptor binding site [18]. A meta-analysis of the correlation between influenza virus egg adaptation and vaccine effectiveness showed that egg adaptation has a strong negative correlation with vaccine effectiveness [19]. In other words, passage adaptation in eggs was found to be a strong contribution to the low vaccine effectiveness against H3N2 [20].
Unlike egg, it is known that antigenic mutations do not occur when influenza virus is cultured in animal cells [212223]. As such, the development of influenza vaccines production systems using Vero or MDCK cells has emerged as a way to resolve the problems pertaining to the egg adaption, and several cell culture influenza vaccines are currently available in the market. Also there are several reports indicating that cell culture influenza vaccine actually showed greater effectiveness than that of egg based vaccine [242526]. Therefore, it is expected that there will be an increasing interest in cell culture-based production of CVVs and influenza vaccine in the future.
In this study, H3N2 virus was incubated 15 times in animal cells or eggs to confirm and compare the presence of amino acid mutations in hemagglutinin and neuraminidase proteins. Prototype H3N2 virus (A/Texas/50/2012) with no history of subculture in eggs was obtained from the Victorian Infectious Diseases Reference Laboratory (VIDRL) for this study and this virus was propagated using both specific pathogen free (SPF) egg and MDCK-Sky3851 cell line which is the qualified cell lines used in cell culture influenza vaccine production, and the presence or absence of mutation in the antigen sequence of the influenza virus collected at each passage was confirmed.
Materials and Methods
Virus, embryonated chicken eggs and cells
Prototype A/Texas/50/2012 (H3N2) influenza virus was obtained from VIDRL (Melbourne, Australia). SPF embryonated chicken eggs were obtained from Namduk SPF (Icheon, Korea) and incubated for 9–10 days at 37℃ prior to virus inoculation. MDCK-Sky3851 cell line, developed and used in influenza vaccine manufacturing by SK Bioscience (Seongnam, Korea), were maintained in culture medium without serum at 37℃, 5% CO2.
Propagation and passage culture of viruses in MDCK-Sky3851 cells and eggs
Virus seed stocks for A/Texas/50/2012(H3N2) influenza virus were prepared through propagation of prototype virus in MDCK-Sky3851 cell line several times and these stocks were used in this study. Virus seed stock was inoculated into MDCK-Sky3851 cells in spinner flask with two different dilution factors of 107–109 and incubated at 34℃, 5% CO2, and 80 rpm agitation. After 2 days, HA titer was checked and the virus showing the highest HA titer was harvested as a viral stock and stored at −70℃. Then, this viral stock was inoculated to MDCK-Sky3851 cell line again and this process was continued up to 15 passages. Virus seed stock was also inoculated into eggs with three different dilution factors of 102–104 and incubated at 37℃ for 3 days followed by collecting the allantoic fluid after storing at 4℃ for overnight. The highest HA titer virus was harvested as an egg passage virus stock and stored at −70℃. This egg virus stock was also inoculated into eggs again in order to make next passage stock and this process was repeated up to 15 passages.
Hemagglutination assay
Samples were 2-fold diluted in v-bottomed 96-well microplates and chicken red blood cells at a concentration of 0.5% were added to each well, and the plates were incubated for 40–50 minutes at room temperature. When the 0.75% guinea pig red blood cells were used, samples were diluted in round-bottomed 96-well microplates and incubate for 1 hour at room temperature. The sedimentation of red blood cells was evaluated and HA titers were reported as hemagglutinin units (HAU) /50 µL.
Gene sequencing and protein structure analysis
Viral RNA was extracted from each cell and egg passage viruses by using extraction kit (Invitrogen, Carlsbard, CA, USA). Reverse transcription polymerase chain reaction was performed with suitable primer to obtain DNA of hemagglutinin and neuraminidase. Nucleotide sequencing analysis was conducted by Macrogen Inc. (Seoul, Korea) and Geneious software (ver. 10.2.3; Biomatters Ltd., Auckland, New Zealand) was used for data analysis and comparison. Protein structure analysis was conducted by using Phyre2 (ver. 2.0; Structural Bioinformatics Group at Imperial College, London, UK) and UCSF-Chimera software (Resource for Biocomputing, Visualization, and Informatics, San Francisco, CA, USA).
Results
HA titer for each passage viruses in cells and eggs
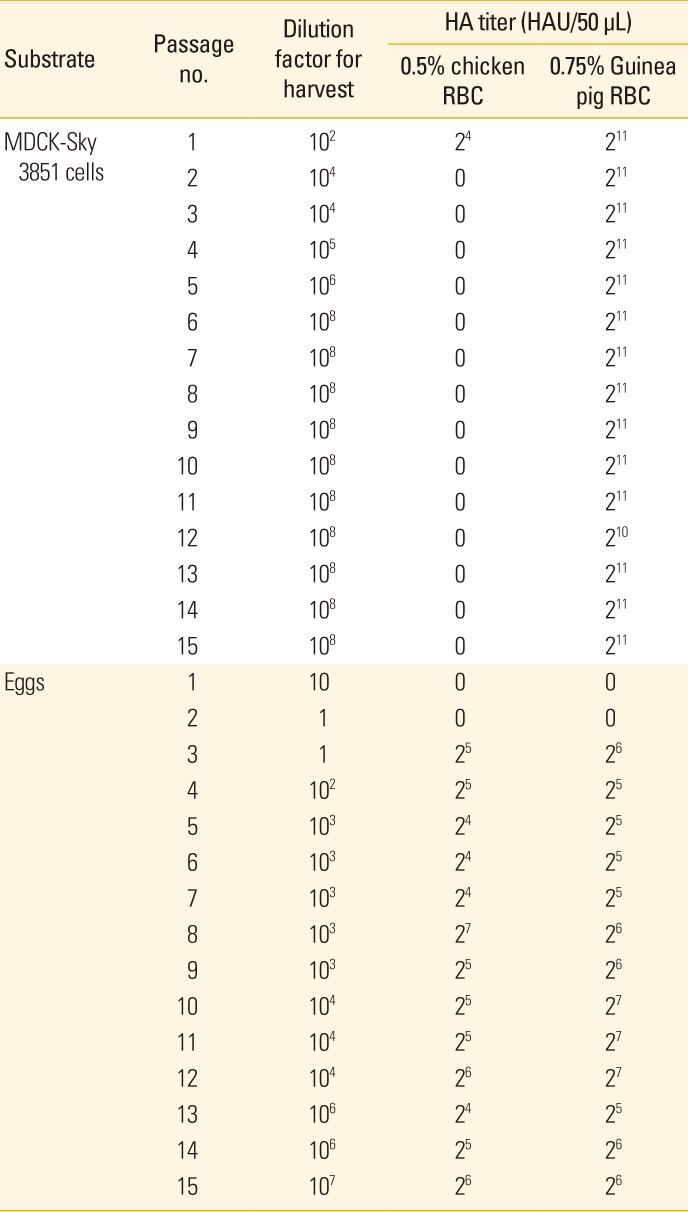
A total of 15 passages in MDCK-Sky3851 cells and eggs were carried out using the prototype A/Texas/50/2012 virus distributed by VIDRL and HA assay was conducted to check the HA titer for viruses of each passage. In case of MDCK-Sky3851 cells, when the chicken red blood cells were used in HA assay, HA titer was identified only in the 1st passage virus (24 HAU/50 µL) and no titers were showed in all other viruses from the 2nd passage. In contrast, when guinea pig red blood cells were used for HA assay, overall HA titer were maintained at 210–211 HAU/50 µL with no significant differences among the 1st to 15th passages and no increase in HA titer was also observed as the passage progressed (Table 1). Unlike in the MDCK-Sky3851 cells, viruses passaged in eggs showed reactivity in both red blood cells up to the 15th passage. Slightly higher HA titers were found in guinea pig red blood cells compared to those of chicken red blood cells from the 3rd passage to 15th passage, 24–27 HAU/50 µL in chicken red blood cells and 25–<28 HAU/50 µL in guinea pig red blood cells. HA titers were not found in the 1st–2nd passage viruses in both erythrocytes (Table 1).
Gene sequence analysis for hemagglutinin and neuraminidase of influenza viruses
The hemagglutinin and neuraminidase gene sequences of each passaged virus were compared with those of parent virus. For the sequence of parent viruses, the sequence (ID no. EPI_ISL_122006) registered in Global Initiative on Sharing All Influenza Data was referred to. In the result of sequencing analysis, it was confirmed that hemagglutinin and neuraminidase sequences were 100% identical to those of parent virus in all passages through the 1st to 15th in MDCK-Sky3851 cells (Table 2). In case of viruses passaged in eggs, hemagglutinin sequence of the 1st to 2nd passage viruses was the same as that of the parent virus, but there were three mutations identified from the 3rd to 8th passage viruses, A515G, G604A, and C942T, and from the 9th passaged virus, additional hemagglutinin sequence mutation with C704T were detected (Table 2). For the neuraminidase sequence, it was confirmed that the 3rd to 15th passage viruses all match 100% of the sequence of the parent virus.
Protein structure analysis of hemagglutinin
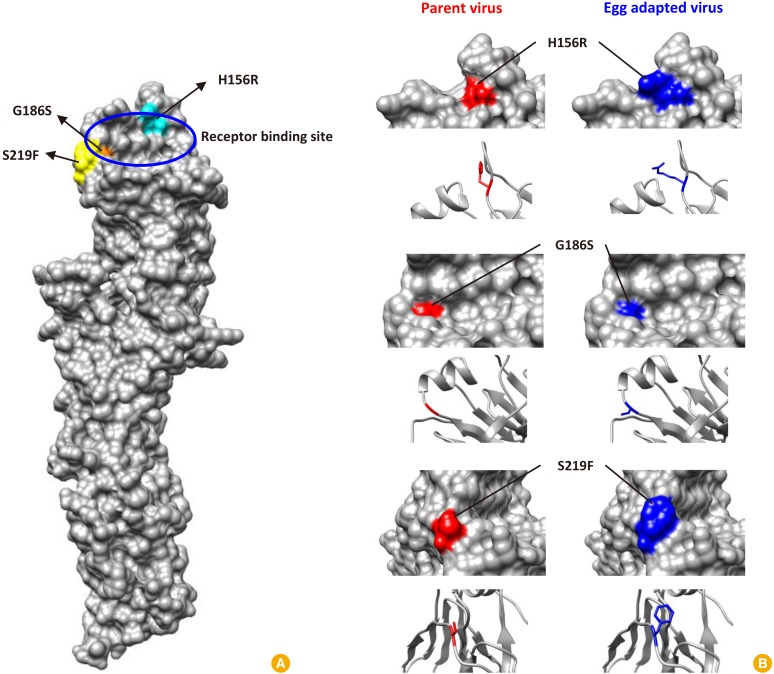
In the gene sequence analysis, four mutations were identified in hemagglutinin during 15 passage culture in eggs and the effect of these mutations on the amino acid sequences was examined. As a result of codon sequence analysis, it was confirmed that three out of four changes in nucleotide sequence resulted in changes in the amino acid sequence and the other one showed silent mutation (Table 2). In order to determine where these amino acid mutations are located and how these structurally affect the receptor binding sites of the hemagglutinin antigen, the structure analysis was performed using Phyre2 software. The analysis revealed that the G186S mutation is relatively inward of the hemagglutinin, while the H156R and S219F mutations are located near the surface of receptor binding site and may cause structural changes (Fig. 1).
Discussion
Mismatch between circulating influenza viruses and vaccine strains cause a reduction of vaccine effectiveness and this phenomenon is known to be associated with the adaptation in eggs during the manufacturing process of the reassortant CVVs [2728]. When the influenza virus is cultured in eggs, antigenic mutations may be caused by egg adaptation, and it is reported that this phenomenon is particularly prominent in H3N2 virus [36]. In this study, in order to confirm the mutations due to egg adaptation of influenza virus, prototype A/Texas/50/2012 virus that had not been passaged in eggs were obtained from VIDRL and the virus was passaged up to 15 times in MDCK-Sky3851 cells, which are Skycellflu vaccine producing cell lines, and in embryonated eggs, respectively, and then HA titer and gene sequences of hemagglutinin and neuraminidase were compared.
The HA titer of the prototype virus prior to the 1st passage is 24 HAU/50 µL for both chicken and guinea pig red blood cells, and when this virus was passaged in MDCK-Sky3851 cells, it was confirmed that the HA titer increased up to 211 HAU/50 µL from the first passage in the guinea pig red blood cells which corresponding to 128 times increase compared to the prototype virus and this was maintained up to the 15th passages. However, HA titer was not detected from the 2nd passage in chicken red blood cells, and it was confirmed that the reactivity of virus may be lowered with chicken red blood cells when passaged in MDCK-Sky3851 cells. In case of egg culture virus, HA titer was not determined in both chicken and guinea pig red blood cells until the 2nd passage, and HA titer began to appear at 25 HAU/50 µL in chicken red blood cells and 26 HAU/50 µL in guinea pig red blood cells from the 3rd passage. Until the 15th passages, HA titers were maintained quite constantly, not showing much variability, with a maximum of 27 HAU/50 µL in chicken red blood cells and a maximum of 28 HAU/50 µL in guinea pig red blood cells. Based on the HA assay results using chicken and guinea pig red blood cells, it was confirmed that the binding avidity of influenza virus varies among the different types of red blood cells depending upon the host species and the substrate for passage of influenza virus. Since the prototype virus used in this study was originated from human, it would be highly reactive to α2,6-linked receptors. Thus, when the virus was cultured in MDCK-Sky3851 cells, the virus was responsive to guinea pig red blood cells which contain α2,6-linked receptors as it is of the same mammalian origin, but not to chicken red blood cells with the α2,3-linked receptors as it is of avian origin. In contrast, when the prototype virus was cultured in egg, it did not grow well with no confirmed HA titer until the 2nd passages since it was not the host species from which the virus had been originally propagated. From the 3rd passage, it seems that the virus became capable to respond to both types of chicken and guinea pig red blood cells as they propagated and adapted to the egg. This suggests that the virus has binding avidity for both α2,3 and α2,6-linked receptors. However HA titer with guinea pig red blood cells was lower for egg cultured virus, compared to that of MDCK-Sky3851 cell cultured virus, suggesting that it may be owing to the egg adaptation.
Subsequently, the nucleotide sequences of the hemagglutinin and neuraminidase genes, which are antigens located on the surface of virus, were analyzed to determine if the antigenicity of the viruses passaged 15 times in MDCK-Sky3851 cells and eggs was altered. As a result, it was confirmed that the nucleotide sequences of the viruses from MDCK-Sky3851 cells were 100% identical to those of the parent virus across all passages from the 1st to 15th without any mutation on both hemagglutinin and neuraminidase sequences. In the case of egg cultured viruses, no mutation was observed in the sequence of the hemagglutinin gene up to the 2nd passage, but there were three changes identified for A516G, G604A, and C942T in the 3rd to 8th passaged viruses, and the corresponding amino acid sequence mutations were H156R, G186S, and silent mutation, respectively. In addition, the C704T mutation was further identified in the 9th to 15th passaged viruses, and the corresponding amino acid sequence mutation was identified as S219F. It was confirmed that these mutations are identical to the amino acid positions (156, 186, 219) that caused antigenic mutations in IVR-165, the H3N2 vaccine strain of the 2012–2013 Northern Hemisphere season, which had low vaccine effectiveness due to antigenic mismatch between the circulating virus and vaccine strain [11]. In the result of neuraminidase gene sequencing analysis of egg cultured viruses, it was confirmed that the 3rd to 15th passages were 100% identical in sequence to those of the parent virus, except for the 1st and 2nd passages which were not sequenced. As the 1st and 2nd passaged viruses had HA titer of 0 HAU/50 µL, the sequence of neuraminidase could not be confirmed by polymerase chain reaction due to low virus contents. However, since no mutations were identified in the neuraminidase gene sequences of the 3rd to 15th passaged viruses, it is expected that the neuraminidase gene sequences of the 1st and 2nd stocks also may not carry any mutations.
In the viruses cultured in egg, a total of three amino acid mutations were confirmed, and a structural analysis was performed to confirm what structural and functional changes these may induce in hemagglutinin antigen. Influenza hemagglutinin has five antigenic sites [29303132], among which antigenic site A and B are located at the distal end from the virus surface and contain receptor binding sites, in particular antigenic site B is known as the immonodominant [33]. The hemagglutinin receptor binding site is composed of 130-loop, 150-loop, 190-helix, and 220-loop structures, forming a concave shape, and the sialic acid is bound here [34]. Influenza viruses have evolved naturally through mutations in receptor binding sites to evade antibody responses from the host. Therefore, amino acid mutations in the position that constitutes the receptor binding site can lead to changes in viral antigenicity through structural changes. In this study, three amino acid mutations were identified in the influenza virus cultured in egg, including H156R, G186S, and S219F. In terms of amino acid position, all of the amino acid mutations were very close to the position of the structure constituting the hemagglutinin receptor binding site, so it can be seen that there is a possibility of inducing changes in receptor binding avidity. According to the structural analysis of the hemagglutinin, the structure before and after mutation was quite different at position 156 where histidine was changed to arginine and 219 where serine was changed to phenylalanine. On the other hand, the change of glutamic acid to serine at amino acid position 186 was not significant in the structural difference (Fig. 1). G186S and H156R are mutations that normally occur characteristically in egg adaptation, and show the effect of improving replication by increasing the binding avidity of α2,3-linked receptors rather than α2,6-linked receptors. However, G186S is known to show an insignificant effect in terms of antigenicity [335]. S219F mutation also affects binding avidity, especially in the case of phenylalanine, which is an amino acid containing a bulky aromatic ring, which may affect 220-loop conformation and physically alter the receptor binding site [3]. This can lead to changes in antigenicity, and it is also known to act synergistically to change the antigenicity of influenza virus, especially when positions 186 and 219 change simultaneously [3]. Thus, all three amino acid mutations identified in this study were related to egg adaptation and were identified as the same positions as amino acid mutations known to affect antigenic changes which result in low vaccine effectiveness of influenza viruses. In addition to these results, it is necessary to conduct animal test to determine whether amino acid mutations are directly related to actual antigenic changes. It would be desirable to confirm the correlation between the incidence of mutation and the actual vaccine efficacy through animal immunization and challenge test using the mutated virus in egg and the non-mutated virus in cell.
In conclusion, it was confirmed that when the H3N2 influenza virus was cultured in egg, amino acid mutations occurred near the receptor binding site of hemagglutinin, which may alter binding avidity and antigenicity accordingly. On the other hand, it was also confirmed that such amino acid mutations did not occur at all when the influenza virus is cultured in MDCK-Sky3851 cells. Therefore, cell culture influenza vaccines production system is a more efficient and reliable method for producing vaccines in that influenza vaccines can be produced without the risk of antigenic changes from the actual prevalent influenza virus. In this respect, the importance of the cell culture influenza vaccine is anticipated to increase further, and it is essential to continuously monitor and evaluate the change of vaccine effectiveness due to amino acid mutation in egg cultured vaccines.
 XML Download
XML Download