

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Protein arginine methylation, which is one of the most common post-translational modifications of proteins in mammalian cells [1], is mediated by the enzymes of the protein arginine methyltransferase (PRMT) family [2]. By methylating various protein substrates, PRMT family members play diverse roles in gene expression, RNA splicing, signal transduction and other processes [3456]. Among the nine isoforms of PRMT, PRMT1 is the major isoform and accounts for approximately 85% of methylarginines in mammalian cells [7]. PRMT1 is ubiquitously expressed in most cells and, unsurprisingly, Prmt1 whole body knock-out (KO) mice are embryonically lethal [89]. Recently, cell type-specific functions of PRMT1 have been investigated using conditional KO mouse models. For example, deletion of Prmt1 from neural progenitors using Nestin-Cre resulted in brain demyelination due to defective development of oligodendrocytes [10], while deletion of Prmt1 from myogenic precursors using Pax7-CreERT2 resulted in the failure of muscle differentiation [11]. Thus, PRMT1 is thought play important roles in the development of different organs.
The pancreas is the central organ responsible for regulating whole-body glucose homeostasis and metabolism by secreting diverse hormones and digestive enzymes [12]. Therefore, proper development of pancreatic endocrine and exocrine cells is essential for the maintenance of normal glycemic control. In the mouse, pancreatic specification initiates at embryonic day 8.5 (E8.5) from pancreatic and duodenal homeobox 1 (PDX1)-positive pancreatic progenitor cells [13]. At E9.5, pancreatic budding and branching morphogenesis occur. At E14.5, the so-called secondary transition occurs, during which pancreatic progenitor cells rapidly proliferate and differentiate [14]. After E14.5, individual pancreatic lineages undergo further differentiation, expansion, and organization [14]. However, although many studies have examined pancreas development, the precise regulatory mechanisms remain unknown. Here, we aimed to study the role of PRMT1 in pancreas development.
METHODS
Mouse experiments
Prmt1 floxed (Prmt1fl/fl) (MGI: 4432476) [15] mice were crossed with Pdx1-Creearly (MGI: 2684317) [16] mice to generate pancreatic progenitor cell-specific Prmt1 knock-out (Pdx1-Creearly; Prmt1fl/fl, herein called Prmt1 PKO) mice. Mice were housed in climate-controlled, specific pathogen-free barrier facilities under a 12-hour light/dark cycle, and chow and water were provided ad libitum. Noon on the morning of vaginal plug discovery was considered E0.5. The animal experiment protocols for this study were approved by the Institutional Animal Care and Use Committee of the Korea Advanced Institute of Science and Technology (KA2011-29). All experiments were performed in accordance with the relevant guidelines and regulations. Body weight and random blood glucose levels of mice were measured in the daytime, with the latter assessed using a Gluco Dr. Plus glucometer (Allmedicus, Anyang, Korea).
Pancreatic tissue preparation
The pancreata of mice were fixed with 4% paraformaldehyde for 2 to 4 hours at 4℃ and washed for 30 minutes with phosphate buffered saline (PBS) at 4℃. Pancreatic tissues were processed with an automatic tissue processor (TP1020; Leica Biosystems, Wetzlar, Germany) and embedded in molten paraffin wax. Paraffin-embedded tissue sections were sliced at a thickness of 4 µm and mounted on adhesive glass slides (081000; Marienfeld, Lauda-Königshofen, Germany).
Hematoxylin and eosin staining
Formalin-fixed paraffin-embedded pancreatic tissue slides were deparaffinized and rehydrated. Hematoxylin and eosin staining was performed as previously described with slight modification [17]. Images were acquired using a bright-field microscope (DS-Ri2 camera; Nikon, Tokyo, Japan) and analyses were performed with the NIS-Elements BR (Nikon) software.
Immunofluorescence staining
Formalin-fixed paraffin-embedded pancreatic tissue slides were deparaffinized and rehydrated. Antigen retrieval was performed by incubating the slides in sodium citrate buffer (10 mM sodium citrate, pH 6.0) for 20 minutes at 95℃. The slides were cooled for 10 minutes at room temperature and washed in PBS for 10 minutes, and the samples were blocked with 4% normal goat serum (005-000-121; Jackson ImmunoResearch, West Grove, PA, USA) in PBS for 1 hour at room temperature. The samples were then incubated for 18 hours at 4℃ with primary antibodies against the following: insulin (A0564, 1:1,000; Dako, Carpinteria, CA, USA), PRMT1 (84361, 1:1,000; Abcam, Cambridge, MA, USA), glucagon (G2654, 1:1,000; Sigma-Aldrich, St. Louis, MO, USA), amylase (A8273, 1:200; Sigma-Aldrich,), mucin 1 (MUC1, MA5-11202, 1:1,000; Invitrogen, Carlsbad, CA, USA), PDX1 (F6A11, 1:500, Developmental Studies Hybridoma Bank [DSHB], Iowa City, IA, USA; 47267, 1:1,000, Abcam), SRY-box 9 (SOX9, AB5535, 1:1,000; Merck Millipore, Burlington, MA, USA), neurogenin 3 (NGN3, F25A1B3, 1:500; DSHB), NK2 homeobox 2 (NKX2.2, 74.5A5, 1:500; DSHB), NKX6.1 (F65A2, 1:500; DSHB), ISL LIM homeobox 1 (ISL1, 40.2D6, 1:500; DSHB), and phosphorylated histone H3 (06-570, Merck Millipore, 1:1,000). The samples were washed in PBS for 10 minutes and incubated for 2 hours at room temperature with the following secondary antibodies: Alexa Fluor 647-conjugated anti-hamster immunoglobulin G (IgG, 127-605-160, 1:1,000; Jackson ImmunoResearch), Alexa Fluor 647-conjugated anti-guinea pig IgG (106-605-003, 1:1,000; Jackson ImmunoResearch), Alexa Fluor 488-conjugated anti-guinea pig IgG (106-545-003, 1:1,000; Jackson ImmunoResearch), Alexa Fluor 488-conjugated anti-rabbit IgG (111-545-144, 1:1,000; Jackson ImmunoResearch), Alexa Fluor 594-conjugated anti-rabbit IgG (111-585-144, 1:1,000; Jackson ImmunoResearch), Alexa Fluor 488-conjugated anti-mouse IgG (115-545-166, 1:1,000; Jackson ImmunoResearch), and Alexa Fluor 594-conjugated anti-mouse IgG (115-585-166, 1:1,000; Jackson ImmunoResearch). The samples were washed in PBS for 10 minutes, incubated for 5 minutes with 4′,6-diamidino-2-phenylindole (DAPI, D9542, 1 µg/mL; Sigma-Aldrich) at room temperature, and then mounted with fluorescence mounting medium (S3023; Dako). Images were acquired using a fluorescence microscope (DS-Ri2 camera; Nikon) and a confocal microscope (LSM 780; Carl Zeiss, Oberkochen, Germany). Imaging analyses were performed using the NIS-Elements BR (Nikon) and ZEN (Carl Zeiss) software packages.
Quantitative reverse transcription polymerase chain reaction
Total RNA was extracted from dorsal pancreas tissues using the TRIzol reagent (15596026; Invitrogen) according to the manufacturer's protocol. Genomic DNA was removed using a TURBO DNA-free kit (AM1907; Invitrogen) and 1 µg of total RNA was used to generate complementary DNA (cDNA) with a High-Capacity cDNA Reverse Transcription kit (4368813; Applied Biosystems, Waltham, MA, USA). Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed with the Fast SYBR Green Master Mix (4385614; Applied Biosystems) and a Viia 7 Real-time PCR System (Applied Biosystems) according to the manufacturer's instructions. Relative Neurog3 expression was analyzed using the delta Ct (threshold cycle) method [18], with the Rplp0 (36B4) detected as a reference gene. The following primers were used to analyze gene expression levels: Rplp0 forward (GAGGAATCAGATGAGGATATGGGA), Rplp0 reverse (AAGCAGGCTGACTTGGT-TGC), Neurog3 forward (CAGTCACCCACTT-CTGCTTC), and Neurog3 reverse (GAGTCGGGAGAACTAGGATG).
Immunoblotting
Dorsal pancreas tissues obtained from mouse embryos at E14.5 were lysed with lysis buffer (10 mM Tris-Cl, 66 mM EDTA, 150 mM NaCl, 0.4% sodium deoxycholate, and 1% NP-40) for 1 hour. The samples were boiled at 100℃ for 5 minutes in sample buffer (100 mM Tris-Cl, 4% SDS, 0.2% bromophenol blue, 20% glycerol, and 200 mM β-mercaptoethanol), centrifuged for 5 minutes at 5,900 G, separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene fluoride membrane (Merck Millipore). The membrane was blocked with 5% dry milk, 0.1% Tween-20 in PBS for 1 hour and blotted with primary antibodies against PRMT1 (84361, 1:1,000; Abcam), NGN3 (F25A1B3, 1:1,000; DSHB) and α-tubulin (sc-8035, 1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) in blocking solution for 18 hours at 4℃. The blot was then incubated with horseradish peroxidase-conjugated secondary antibodies (Cell Signaling Technology, Danvers, MA, USA) for 1 hour, and proteins were visualized using a ChemiDoc System (Bio-Rad, Hercules, CA, USA).
Protein degradation assay
Adenoviruses expressing nonspecific RNAi or Prmt1 RNAi [15] were transduced into cells 24 hours before transfection. Hemagglutinin (HA)-NGN3 plasmids were transfected into 30% to 50% confluent mPAC cells in a 6-well plate using cationic lipids (FuGene 6; Roche, Basel, Switzerland) according to the manufacturer's protocol. Additional protein synthesis was blocked 48 hours after transfection by the addition of cycloheximide (CHX) at 100 µg/mL. Cells were lysed with RIPA buffer at 0, 7.5, 15, 30, 60, and 90 minutes after CHX treatment.
Quantification analysis
Cell counting and β-cell area measurements were done using the ImageJ (National Institutes of Health) program. For quantitative analysis, pancreata of age-matched wild type (WT) littermates and Prmt1 PKO mice were whole-sectioned at 4 µm. Every fifth (20 µm apart) sections was selected for immunofluorescence staining. The β-cell area was calculated by dividing the insulin-positive area by the total pancreatic area.
Statistical analysis
All values are expressed as the mean±standard error of mean. The two-tailed Student's t-test or one-way analysis of variance (ANOVA) followed by post hoc Bonferroni's test were used to compare groups. P values below 0.05 were considered statistically significant. The levels of significance indicated in the graphs are P<0.05, P<0.01, and P<0.001.
RESULTS
Defective pancreas development in Prmt1 PKO mice
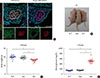
To explore the functional role of PRMT1 in pancreas development, we generated Prmt1 PKO mice by crossing Prmt1fl/fl mice with Pdx1-Creearly mice. Prmt1 PKO was confirmed by immunofluorescence staining in embryonic pancreas at E10.5, which showed that there was no detectable level of PRMT1 in PDX1-positive pancreatic progenitor cells (Fig. 1A). Prmt1 PKO mice were born normally at the expected Mendelian ratio. However, Prmt1 PKO mice exhibited growth retardation, a lower body weight and severe hyperglycemia, with a blood glucose level greater than 500 mg/dL at 4 weeks of age (Fig. 1B–D). In contrast, heterozygous Prmt1 PKO (Pdx1-Creearly; Prmt1fl/+) mice were normal in their body weight and blood glucose level.
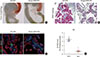
Gross morphological analysis at postnatal day 7 (P7) showed that Prmt1 PKO mice were characterized by a severely hypoplastic pancreas (Fig. 2A). Further histological analysis with H&E staining revealed that the numbers of acini, ductal cells and islet cells were robustly reduced in Prmt1 PKO pancreas at P0, and that the Prmt1 PKO pancreas was mostly composed of progenitor epithelial cells (Fig. 2B). Immunofluorescence staining using anti-insulin and anti-amylase antibodies revealed that the numbers of acinar cells and β-cells were reduced in Prmt1 PKO pancreas (Fig. 2C). Further quantitative analysis confirmed that the β-cell area was significantly reduced in the pancreata of Prmt1 PKO mice (Fig. 2D). In addition, co-immunostaining for the ductal epithelial cell marker, MUC1, plus insulin showed that the progenitor epithelial plexus, which normally disappears during the late embryonic period, was still evident in the Prmt1 PKO pancreas at P0 (Fig. 2C). Thus, pancreas development, including both exocrine and endocrine cells, was severely compromised in Prmt1 PKO mice. These data indicate that PRMT1 is required for the normal development of the pancreas.
PRMT1 is required for endocrine cell commitment during the secondary transition
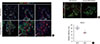
To investigate the underlying cause of the hypoplastic pancreas seen in Prmt1 PKO mice, we carefully observed their pancreas development. Until E13.5, Prmt1 PKO mice showed normal pancreas development, with normal expression of pancreas development markers, such as NKX2.2, PDX1, SOX9, and NGN3 (Fig. 3A). However, at E14.5, severe defects in endocrine development were observed in Prmt1 PKO mice, with robust decreases in the numbers of cells that expressed the endocrine cell differentiation markers, NKX2.2, NKX6.1, and ISL1 (Fig. 3B). These data indicate that PRMT1 plays an important role in endocrine cell differentiation during the secondary transition.
Prmt1 PKO mice exhibit increased numbers of NGN3-expressing cells during the secondary transition
The secondary transition, which is a critical step in pancreas development, is characterized by neogenesis and differentiation of endocrine cells, as well as active proliferation of progenitor cells [19]. During this stage, transient expression of NGN3 plays a central role in regulating the formation of endocrine cells [202122]. Sustained expression of NGN3 in pancreatic progenitor cells results in premature endocrine cell differentiation and defects in exocrine cell development [23]. These previous data motivated us to test NGN3 expression in Prmt1 PKO mice. Notably, we found that Prmt1 PKO mice showed a robust increase in the number of NGN3-positive cells that remained in the SOX9-positive trunk epithelium at E14.5 (Fig. 4A). Immunofluorescence staining of NGN3, SOX9, and MUC1 indicated that the NGN3-positive cells in Prmt1 PKO mice failed to delaminate and further differentiate into endocrine cells (Fig. 4A). During the secondary transition, NGN3-positive cells are well known to upregulate the Notch signaling of surrounding cells to induce proliferation [24]. Interestingly, co-immunostaining of the proliferation marker, phosphorylated histone H3, plus PDX1 showed that the proliferation of pancreatic progenitors was significantly reduced in Prmt1 PKO mice compared to WT mice (Fig. 4B and C). These data indicate that PRMT1 is necessary for proper NGN3 expression in pancreatic cells during the secondary transition, which is crucial for normal development of the pancreas.
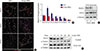
PRMT1 is necessary for rapid degradation of the NGN3 protein
During pancreas development in the mouse, NGN3 expression peaks at E14.5-15.5 and rapidly decreases within 24 hours [25]. However, we observed prolonged expression of NGN3 until E18.5 in the pancreas of Prmt1 PKO mice (Fig. 5A). To investigate the mechanism underlying this prolonged expression of NGN3 in the pancreas of Prmt1 PKO mice, we examined Neurog3 gene expression by qRT-PCR and NGN3 protein expression by Western blot analysis. As the mRNA and protein expressions of NGN3 were unaltered in the pancreas of Prmt1 PKO mice (Fig. 5B and C), we hypothesized that PRMT1 may be involved in altering the stability of the NGN3 protein. To test this hypothesis, we performed protein degradation assays in mPAC cells exposed to CHX, which blocks the translational elongation step of protein synthesis. Interestingly, knock-down of Prmt1 in mPAC cells was found to trigger defects in NGN3 protein degradation (Fig. 5D). These results indicate that PRMT1 is required for the rapid degradation of the NGN3 protein, but does not alter its protein expression.
DISCUSSION
In this study, we demonstrated the functional role of PRMT1 in pancreas development. Prmt1 PKO mice showed severe hyperglycemia with a hypoplastic pancreas. In these mice, developmental defects of the pancreas were observed beginning at the secondary transition. Notably, NGN3-expressing cells were robustly increased in the pancreas of Prmt1 PKO mice from E14.5, and this was due to failure of the rapid degradation of NGN3 that was seen in WT mice. Indeed, our results confirmed that PRMT1 is necessary for rapid degradation of the NGN3 protein.
Pancreas development comprises sophisticated cascades of transcription factor expressions that directs the differentiation and proliferation of individual pancreatic cell types [19]. Here, we show that PRMT1 is required for the transient expression of NGN3 during the secondary transition. However, it is still unclear how the observed increase in NGN3-positive cells translates to the severe hypoplastic phenotype of the pancreas in Prmt1 PKO mice. Based on the expression level and spatial localization of NGN3, individual NGN3-positive cells can differ in their developmental fate and lineage commitment [2627]. Therefore, precise time course-observations of NGN3-positive cells by lineage-tracing experiments are necessary in Prmt1 PKO mice.
Further studies are also needed to uncover the detailed mechanism through which PRMT1 regulates NGN3 protein stability. Recent work has shown that cyclin dependent kinase (CDK) induces the multiple phosphorylation of NGN3, thereby promoting its protein degradation via the ubiquitin-proteasome pathway [2829]. Therefore, PRMT1 may destabilize the NGN3 protein either directly via methylation or indirectly via enhancement of phosphorylation.
Since PRMT1 has a broad substrate specificity, it may play NGN3-independent roles in pancreas development may exist. Through its well-known function as a transcriptional coactivator, PRMT1 potentiates gene expression levels by recruitment of coactivator associated arginine methyltransferase 1 (CARM1) (PRMT4) or methylation of histone H4 [330]. In this manner, PRMT1 can directly turn on gene subsets that are related to pancreatic endocrine and exocrine cell development. On the other hand, PRMT1 modifies the activities of signaling proteins via direct methylation to regulate pathways that are essential for tissue development, such as the Wnt/β-catenin, transforming growth factor β and Notch signaling pathways [53132]. Collectively, these effects plus the up-regulation of NGN3-expressing cells may largely account for the phenotype of Prmt1 PKO mice.
In conclusion, we herein show that PRMT1 plays an essential role in pancreas development by regulating NGN3 protein stability. Our work provides a novel mechanistic insight into pancreas development and may inform islet regeneration studies, and thus has potential implications for the treatment of diabetes.

 XML Download
XML Download