

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Impaired endothelial function is a fundamental component of the pathogenesis of cardiovascular disease. The endothelium plays an important role in maintaining vascular health via synthesis of various vasoactive mediators such as nitric oxide (NO), prostacyclin and endothelium-derived hyperpolarizing factors [1]. Exercise training is generally known to improve endothelial function [2345]. This beneficial effect of exercise training has been associated with increased expression of endothelial nitric oxide synthase (eNOS) [6], enhanced production and bioavailability of NO [7] and improved endothelium-dependent relaxation (EDR) [8]. However, these effects are inconsistent. For instance, Green et al. [9] reported non-uniform changes in flow-mediated dilation (FMD), a surrogate for endothelial function, in response to exercise training, including individuals who had no or negative changes in endothelial function following exercise training. These authors also noted that greater changes in endothelial function after exercise training were often observed in individuals having lower pre-training cardiopulmonary fitness level and baseline endothelial function, suggesting an inverse relationship between initial level and trainability for endothelial adaptation.
Accumulating data suggest that endothelial function is genetically influenced. We and others showed that EDR is determined by genetic background in mice [101112]. In humans, the estimated heritability of FMD ranges from 0.14 to 0.44 [13141516], suggesting that up to 45% of the variation in FMD is due to genetic factors. Others also reported that changes in FMD in response to training are more correlated in monozygotic than dizygotic twins [1718], implying that endothelial responses to exercise training are also partially affected by genetic factors.
The benefits of exercise training on the cardiovascular system appear to be dependent on the training intensity. Clinical trials and animal studies reported greater cardioprotective effects of high intensity training (75%–90% of VO2max) on VO2max, blood pressure and glucose control compared to moderate intensity training (< 70% of VO2max) [3192021]. Conversely, several studies showed that high intensity training exerts similar effects as moderate intensity training, or no effect, on cardiovascular health [222324]. There is also some evidence linking high intensity exercise to negative outcomes [25262728]. The effects of exercise training intensity on endothelial function are also inconsistent. While greater improvements in endothelial function in response to high intensity training compared to moderate intensity training have been observed in both humans and rats [293031], others have indicated no differences between moderate and high intensity training on endothelial function [332]. These inconsistent results might be due to the heterogeneity in age, sex and baseline health status [32527293132]. However, the contribution of genetic background to these heterogeneous responses has not been formally considered.
Therefore, we aimed to characterize for the first time the genetic contribution to endothelial adaptation to exercise training designed with two different intensities in mouse aortas. Specifically, we examined the effect of two different training intensities (high vs. moderate) on endothelial function in isolated thoracic aortas from four different inbred mouse strains. We hypothesized that all strains would improve endothelial function with training, but the magnitude of improvements would vary based on genetic background and training intensity.
METHODS
Animals
All procedures adhered to the established National Institutes of Health guidelines for the care and use of laboratory animals and were approved by the Institutional Animal Care and Use Committee at Texas A&M University (AUP # 2013-0223). Sevenweek old male mice from four inbred strains, C57BL/6J (B6), 129S1/SvImJ (129S1), SJL/J (SJL), and NON/ShiLtJ (NON), were purchased from Jackson Laboratories. Upon arrival, all mice were given one week to acclimatize to their new environment. These strains were chosen since these four strains are originated from different evolutionary genetic backgrounds [33], and previous studies from our lab and others have demonstrated the differences in intrinsic EDR [101112] and adaptions to exercise training [34]. Mice from each strain were randomly assigned to one of three groups: sedentary (SED, n = 6), exercise training with continuous running at moderate intensity for 4 weeks (MOD, n = 6) or exercise training with interval running at high intensity for 4 weeks (HIT, n = 6). All mice were allowed food and water ad libitum and maintained on a 12:12-h light-dark cycle that initiated at 6:00 AM in the animal housing facility at Texas A&M University. Body weights were collected once a week throughout the study.
Exercise performance test and training
All mice (8-week old) were familiarized to treadmill running (10 min/d) for two days on a six-lane motorized rodent treadmill (Columbus Instruments, Columbus, OH, USA). Each mouse then completed two exercise performance tests separated by 48 h on the treadmill to determine baseline exercise capacity as described previously [35]. Briefly, the treadmill was started at 9 m/min at 0° grade for 9 min as a warm-up. The grade was then increased 5° every 9 min up to a final grade of 15° and speed was increased 2.5 m/min from a starting speed of 10 m/min every three minutes until exhaustion. Exhaustion was defined as an inability to maintain running in spite of repeated contact with the electric grid and manual stimulation. At exhaustion, each mouse was immediately removed from the treadmill and returned to its home cage. Baseline exercise capacity was estimated for each animal using total running time (min). The average for two tests was used to determine the running speed for their training.
HIT mice performed 6 sets of running at 85% of maximal speed for 8 minutes followed by 2 min-active rest at ~50% of maximal speed each session. This HIT protocol was slightly modified from previous studies [321] which reported improvements in endothelial function, cardiac function and endurance exercise capability in rats and/or mice. MOD mice performed continuous running at 65% of maximal speed as described previously [35]. In MOD mice, running time for each strain was varied to approximately match the daily running distance of HIT mice (~70 min a day). All mice were trained 5 days/wk at a 10° incline on the treadmill for 4 weeks at ambient temperature (~24℃). All mice completed all training sessions. After training, two post-training exercise performance tests were conducted as described above.
Tissue harvest and vascular ring experiments
Approximately 48 h after the final exercise bout, mice were weighed and anesthetized by intraperitoneal injection of a cocktail of ketamine (80 mg/kg) and xylazine (5 mg/kg). Thoracic aortas were obtained and prepared for ex vivo vasoreactivity measurement as described previously [12]. Briefly, connective tissue in freshly dissected aortas was carefully removed in ice-cold physiological saline solution (in mmol/l: 118.3 NaCl, 4.7 KCl, 2.5 CaCl2, 1.2 MgSO4, 1.2 KH2PO4, 25 NaHCO3, and 5.5 glucose, pH 7.4) under a microscope, and then aortas were cut into 2 mm segments. Each ring segment was suspended in organ chamber of 610M Multi Chamber Myograph System (Danish Myo Technology, Aarhus, Denmark) filled with 8 ml of oxygenated (95% O2, 5% CO2) physiological saline solution, stretched to the resting tension of 12 mN, and allowed to equilibrate at 37℃ for an hour. This resting tension was determined by the tension-force assessment in response to 25 mM of potassium chloride. Cumulative concentration-response curves to phenylephrine (PE, 10−9 to 10−5 M) and potassium chloride (KCl, 5–100 mM) were generated to assess contractile function, while cumulative concentration-response curves to acetylcholine (ACh) and sodium nitroprusside (SNP) (10−9 to 10−5 M) were generated to assess endothelium-dependent and -independent relaxation, respectively. Concentrationresponse curves to ACh and SNP were assessed after the ring was pre-constricted with 3 × 10−7 M of PE, which generated approximately 70% of maximal constriction in cumulative concentration response curves to PE. Doses were added after the response curve reached a plateau from the previous dose. Unused segments of thoracic aorta were snap-frozen in liquid nitrogen and stored for gene expression profiling analysis. Right gastrocnemius muscles were harvested, cut in half longitudinally and snap-frozen. All collected tissues were stored at −80℃.
Oxidative enzyme activity
Citrate synthase (CS) and succinate dehydrogenase (SDH) enzyme activity were measured as markers for exercise training efficacy. The lateral portion of the right gastrocnemius muscle was placed in 20 volumes of ice-cold sucrose muscle homogenization buffer (20 mM Tris, 40 mM KCl, 2 mM EGTA, 250 mM Sucrose, pH was adjusted to 7.4) and homogenized using the FastPrep-24 (MP Biomedicals, Santa Ana, CA, USA). Homogenates were centrifuged at 600 g for 10 min at 4℃. The supernatants were collected, and total protein concentration was measured with BCA protein assay reagent and pre-diluted BSA standards (ThermoFisher Scientific, Waltham, MA, USA) on Nanodrop 2000 (Thermo-Fisher Scientific). Then citrate synthase and succinate dehydrogenase activity in the protein samples were assessed on the Genesys 10 UV spectrophotometer (ThermoFisher Scientific) following a previously published protocol [36]. Briefly, for CS enzyme activity, 300 µl of distilled water, 500 µl of Tris (200 mM, pH 8.0) with Triton X-100 [0.2%(vol/vol)], 100 µl of 5,5′-dithiobis-(2-nitrobenzoic acid) (1 mM), 30 µl of Acetyl CoA (10 mM) and 20 µl of muscle homogenates were added to a 1-ml cuvette and the baseline activity at 412 nm was measured for 3 min. Then, 50 µl of oxaloacetate (10 mM) was added and the absorbance change at 412 nm was measured for 3 min. For SDH enzyme activity, 661 µl of distilled water, 50 µl potassium phosphate buffer (500 mM, pH 7.5), 20 µl of fatty acid-free bovine serum albumin (50 mg/ml), 30 µl of potassium cyanide (10 mM), 50 µl of succinate (400 mM), 145 µl of 2,6-dichlorophenol indophenol, and 40 µl of muscle homogenates were collected in a 1-ml cuvette and incubated at 37℃ for 8 min. The baseline activity was measured at 600 nm for 3 min. After starting the reaction by adding 4 µl of decylubiquinone (12.5 mM), the absorbance change at 600 nm was recorded for 3 min. The molar extinction coefficients of 13.6 and 19.1 mM−1 cm−1 were used for CS and SDH enzyme activity calculations, respectively. All assays were carried out at room temperature unless specified. CS from porcine heart (Sigma-Aldrich, St. Louis, MO, USA) was used as a standard for CS assay calibration.
RNA isolation and cDNA synthesis
Frozen aortas from B6, SJL, and NON mice were homogenized, and total RNA was isolated utilizing RNeasy fibrous tissue mini kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instruction. Aortas of 129S1 mice were excluded due to the similar vasomotor responses to exercise training as B6 mice (as referred to Figs. 3, 4, 5, 6). The quantity and quality of total RNA were determined by Nanodrop 2000 (ThermoFisher Scientific) and BioAnalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA), respectively. RNA (500 ng) having an integrity number ≥ 7.5 was transcribed into cDNA using RT2 First Strand cDNA Synthesis kit (Qiagen) following the manufacturer's manual. Synthesized cDNA was stored overnight at −20℃.
Gene expression profiling by PCR array
Gene expression profiling specific for mouse endothelial cell biology (Cat. No. PAMM-015Z) was conducted using RT2 profiler PCR array (Qiagen) on a 96-well format. Each plate consists of 84 key genes associated with endothelial cell biology, 5 housekeeping genes, 1 mouse genomic DNA control, 3 reverse transcriptase control, and 3 positive PCR controls. The complete list of genes can be found at the manufacturer's homepage (https://www.qiagen.com/kr/shop/genes-and-pathways/data-analysis-centeroverview-page/). The Real-Time PCR Array was performed as indicated in the user manual with RT2 SYBR Green ROX qPCR Mastermix (Qiagen) using the StepOne Plus (Applied Biosystem, Waltham, MA, USA). Ct values for all genes were normalized with the geometric mean of five housekeeping genes, and the relative gene expression level was determined using the web-based data analysis software provided by Qiagen (https://dataanalysis.qiagen.com/pcr/arrayanalysis.php). With results from PCR array, overrepresentation analysis was conducted in the Ingenuity Pathway Analysis (IPA; Ingenuity Systems, Redwood City, CA, USA; www.Ingenuity.com). Genes differentially expressed between strains and/or groups (p < 0.05) were queried against the pathway gene sets available in the IPA Knowledge Base to identify canonical pathways and molecular/cellular functions in which differentially expressed genes are involved. The core analysis was performed with a specific choice of ‘mouse’ for species and ‘endo thelial cell’ for tissues/cells.
Nitrotyrosine enzyme-linked immunosorbent assay (ELISA)
Because nitrotyrosine is a product of protein tyrosine nitration resulting from oxidative damage to proteins by peroxynitrite, we measured the abundance of nitrotyrosine as an oxidative stress marker in muscle. The medial portion of the gastrocnemius muscle was homogenized and total protein was extracted using Cell Extraction Buffer (Invitrogen, Waltham, MA, USA). Total protein concentration was determined as described above. The abundance of nitrotyrosine in the protein samples was measured using a 3-Nitrotyrosine ELISA kit (Abcam, Cambridge, MA, USA) on the DTX800 Multi-mode microplate reader (Beckman Coulter, Brea, CA, USA) according to the manufacturer's instruction.
Statistical analysis
Values are presented as mean ± standard error (SE). Percent vasocontractile responses (%) were calculated for PE and KCl as [(DP − DB)/DB] X 100, where ‘DP’ is the maximal force generated by a given specific dose and ‘DB’ is the baseline force. Percent vasorelaxation responses for ACh and SNP were calculated as [(DP − DD)/(DP − DB)] X 100, where ‘DP’ is the maximal force pre-generated by PE, ‘DD’ is the lowest force generated at a given dose of ACh or SNP and ‘DB’ is the baseline force. The log10 of half-maximal effective concentration (EC50) for vasoconstriction responses and half-maximal inhibitory concentration (IC50) for vasorelaxation responses were calculated using absolute values (mN) from cumulative concentration-response curves using Prism 6 (GraphPad Software, La Jolla, CA, USA) as the indicator for the sensitivity to a vasoactive agent.
Differences in cumulative concentration-response curves across groups within a strain or among SED groups of 4 inbred mouse strains were compared using One-way ANOVA with repeated measures. Differences in body weight, mitochondrial enzyme activities, EC50 or IC50, relative gene expression, and nitrotyrosine abundance across SED groups of each strain or groups within a strain were analyzed using Two-way ANOVA (strain × group) followed by Tukey post-hoc test to examine the interaction between strain and exercise training mode. Intrinsic differences in the sedentary state between inbred strains were compared to SED of B6. All statistics were performed using JMP 8 (SAS, Cary, NC, USA). Statistical significance was set at p < 0.05.
RESULTS
Table 1 displays differences in pre-training body mass (pre-BM) and exercise capacity, changes in BM and exercise capacity (postminus pre-training) and terminal heart mass (HM) to BM ratio (HM:BM) among strains or groups. Before training, NON had higher and 129S1 and SJL had lower BM than B6. NON and 129S1 SED gained less BM than B6 SED after 4 weeks of the sedentary period. After training, B6 HIT, SJL HIT, and NON MOD showed significantly smaller increases in BM compared with SED of the same strain. A significant interaction (F = 3.58, p < 0.01) between strain and training intensity was identified for the change in BM. HM:BM ratio was similar among groups within the same strain except for NON which had greater HM:BM in both MOD and HIT than their counterpart SED.
As previously found [34], there was a significant strain difference in pre-training exercise capacity, showing that B6 had the lowest and NON had the highest run to exhaustion time. After 4 weeks of sedentary period, NON SED had significantly decreased exercise capacity, while exercise capacity in B6, 129S1 and SJL SED was maintained over 4 weeks of the sedentary period. After training, a significant interaction (F = 3.37, p < 0.01) was found for changes in exercise capacity between strain and training intensity. All four strains of MOD had significant increases in exercise capacity, while for HIT, only NON showed increased exercise capacity compared to their counterpart SED. These data indicate that changes in exercise capacity after training are influenced by the interaction between genetic background and training intensity.
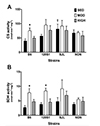
CS and SDH enzyme activities, markers for exercise training efficacy, were not different across SED groups (Fig. 1). In trained mice, MOD had higher CS and/or SDH activity in B6 and 129S1 compared with SED of the same strain. However, all HIT groups had similar CS and SDH activities compared with counterpart SED.
There are differences in intrinsic vasoreactivity across SED mice from four inbred strains (Fig. 2). NON SED had significantly lower EDR compared with B6 SED (Fig. 2A). Conversely, endothelium-independent relaxation was not different among SED groups (Fig. 2B). For intrinsic contractile function, SJL SED had significantly lower contractile responses to both PE and KCl, while 129S1 SED had significantly greater contractile responses to both PE and KCl than B6 SED (Fig. 2C, D). EC50 for KCl varied across strains in SED groups, with SJL SED being less sensitive to KCl relative to B6 SED (Table 2). These results demonstrate the genetic influence on intrinsic vascular function.
The effect of exercise training on aortic relaxation responses is shown in Figs. 3 and 4. Both MOD and HIT of NON had greater EDR compared with NON SED. In contrast, EDR in both exercise-trained groups (MOD, HIT) of B6 and 129S1 were not different from their respective SED. Interestingly, EDR was significantly impaired in SJL HIT compared with SJL SED. Sensitivity (IC50) to ACh was not different across training groups for any of the strains (Table 2). The interaction between strain and training intensity on EDR was significant (F = 1.01, p < 0.01). On the contrary, SNP-induced endothelium-independent vasorelaxation and SNP IC50 were not different among groups within each strain (Fig. 4 and Table 2), indicating that the effect of exercise training on EDR, but not endothelium-independent relaxation, is influenced by the interaction between genetic background and training intensity.
For contractile responses to PE (Fig. 5), there were no differences between SED and exercise groups of B6, 129S1 and SJL. Only NON had decreased contractile responses to PE in both MOD and HIT compared with SED. These decreased responses were accompanied by decreased sensitivities (EC50) to PE (Table 2). Contractile responses to KCl were similar between SED and exercise groups in all strain (Fig. 6). Only SJL MOD had decreased sensitivity (EC50) to KCl compared with SJL SED (Table 2).
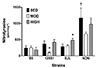
Because of impaired EDR in SJL HIT, we measured the abundance of nitrotyrosine to assess the effect of exercise training on oxidative stress in skeletal muscle (Fig. 7). NON SED had a higher intrinsic level of nitrotyrosine than B6 SED. Nitrotyrosine abundance was lower in 129S1 MOD compared with respective SED. For SJL, nitrotyrosine abundance was higher in HIT than SED. A significant interaction (F = 7.00, p < 0.01) between strain and training intensity was found.
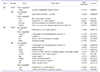
To identify genes differentially regulated by genetic background and/or training intensity, we conducted expression profiling for endothelial cell biology-related genes. The pattern of gene expression changes by exercise training is visualized in Supplementary Fig. 1 and the lists of genes differentially regulated (p < 0.05) are shown in Tables 3 and 4. For B6, only a few genes were differentially regulated in MOD (1 down-regulated) or HIT (1 up- and 4 down-regulated) compared with SED. For SJL mice, 1 gene was up-regulated and 4 genes were down-regulated in MOD, while 9 genes were up-regulated and 2 genes were down-regulated in HIT (Table 3). NON MOD had 3 and 6 genes up- and down-regulated by exercise training, respectively. For NON HIT, 28 genes (20 up-regulated and 8 down-regulated) were differentially expressed (Table 4). There was very little overlap between genes differentially expressed in MOD and HIT within the same strain. Together, these results suggest that the influence of exercise training on transcriptional activation in endothelium is both strain- and training intensity-dependent.
Gene expression profiling data were imported into IPA to identify biological pathways and molecular/cellular functions associated with genes differentially expressed in exercise-trained groups. Only data for SJL HIT, NON MOD, and NON HIT were analyzed because of limited changes in the other groups. IPA identified several canonical pathways and molecular/cellular functions associated with genes differentially regulated by exercise training (Tables 5 and 6). Genes differentially expressed in HIT groups of SJL and NON were overrepresented in pathways related to inflammatory molecule adhesion and migration, while a gene cluster differentially expressed in NON MOD was associated with vessel growth and NO signaling pathways. For the overrepresented molecular and cellular functions, genes differentially expressed in SJL HIT were associated with up-regulation of reactive oxygen species (ROS) production, whereas the effects of HIT in NON were associated with down-regulation of cell adhesion and migration. The genes differentially expressed in NON MOD were related to cell differentiation and proliferation.
DISCUSSION
We evaluated the effect of exercise training with two different intensities on aortic reactivity in four different inbred mouse strains to characterize for the first time the interactive effect between genetic background and training intensity on vasomotor function. The primary findings of this study were: 1) there was a significant interaction between genetic background and training intensity on endothelial responses to exercise training; 2) endothelial gene expression profiles were different depending on both genetic background and training intensity. Supporting the primary outcomes, we also found a significant strain by training intensity interaction for responses of skeletal muscle oxidative enzyme activity and oxidative stress marker to exercise training.
As we previously reported [12], intrinsic vascular function varied among SED groups by mouse strain (Fig. 2), indicating the genetic contribution to intrinsic vascular function in mouse aortas. Compared to B6 SED, NON SED had impaired EDR (Fig. 2A) and also moderately decreased gene expression of eNOS (Fold regulation = −1.48, p = 0.07, data not shown). Additionally, NON SED had a higher skeletal muscle nitrotyrosine in the sedentary state compared with B6 (Fig. 7), suggesting increased baseline oxidative stress in NON. Increased oxidative stress is known to reduce NO bioavailability and subsequent response to endothelium-dependent vasorelaxing agents [3738]. Although our data, as well as previous studies [3940], suggest that NON exhibits phenotypes associated with endothelial dysfunction, the underlying mechanism is not clear and further studies are necessary. For 129S1 and SJL, intrinsic endothelial function was not different from B6. This is in contrast to previous findings that responses to ACh were impaired in aorta from 129P3/J and 129X1/SvJ mice, sub-strains of the 129 strain group [10]. Chen et al. [11] also reported that endothelial function was impaired in aorta from SJL mice relative to C3H/HeJ. These discrepant results could be due to differences in age, sex, sub-strain and/or experimental procedure. For example, Chen et al. [11] used both male and female mice (16- to 18-week old), while only male mice (13-week old) were utilized in this study. SNP-induced endothelium-independent vasorelaxation was similar across inbred strains (Fig. 2B). This result is consistent with previous findings [1011], demonstrating that vascular smooth muscle responses to NO are generally not variable and thus might not be significantly influenced by genetic factors in young animals.
Previous studies have demonstrated that impaired endothelial function is accompanied by exaggerated vasoconstrictor responses [241]. However, in the current study, contractile responses to PE and KCl are greater in 129S1 and lower in SJL than B6 despite having similar EDR. Also, NON, which showed impaired endothelial function, did not show enhanced contractile responses to PE and KCl compared to B6, suggesting that intrinsic contractile function in aortas at least from these four inbred strains is not inversely related to intrinsic endothelial function.
Our results that exercise training had variable effects on vascular function reveal a significant interaction between genetic background and training intensity. For NON, EDR was greater in MOD and HIT than SED (Fig. 3D), accompanied by lower PE-induced vasoconstriction (Fig. 5D). Previous studies reported that exercise training increased expression of eNOS and SOD-1 genes, bioavailability of NO and the release of NO [67]. Neither eNOS nor SOD-1 gene expression were altered by exercise training in NON mice (Table 4). Thus, training-induced improvements in EDR in NON might be due to improvements in other pathways contributing to endothelial-dependent vasorelaxation. For example, prostacyclin synthase (Ptgis) expression was increased in NON MOD. In contrast, markers of oxidative capacity in skeletal muscle were not different among exercise training groups and SED in NON (Fig. 1). Green et al. reported that exercise training-induced change in endothelial function did not correlate with the change in VO2max [9]. In the same sense, our results provide evidence that endothelial responses to exercise training are independent of the change in skeletal muscle oxidative capacity.
There are many factors, such as age, sex, training duration, exercise intensity, and genetic regulation, which can influence the effect of exercise training on endothelial function [394243]. We matched the age and sex of experimental groups to avoid some of these confounding effects. We also chose to train mice for 4 weeks because the training duration of at least 4 weeks has been shown to induce an improvement in aortic endothelial function in healthy animals [43]. However, other studies have reported that endothelial function is not improved by exercise training in young healthy humans and animals [444546]. At least three possible factors might explain this lack of exercise-induced improvements in endothelium-dependent vasorelaxation in B6, 129S1 and SJL. First, our training protocol might exert insufficient stimuli to change endothelial function in aortas from these strains. However, our training volume was sufficient to increase oxidative enzyme activity in skeletal muscle in B6 and 129S1 (Fig. 1). Secondly, structural adaptation could have occurred before post-training measurements in those strains, normalizing the effect of increased vasorelaxant stimuli on vascular smooth muscle relaxation [59]. Unfortunately, we did not assess vessel size in the present study. Thirdly, the intrinsic endothelial phenotype in those strains might be near optimal levels (≥ 75%), thus endothelial function may not be augmentable above the optimal levels (i.e., a ‘ceiling effect’) [4245]. The small number of genes differentially regulated by exercise training supports this concept (Table 3). The one notable exception in B6 HIT is the increased expression of SOD-1, an antioxidant pathway gene that is commonly reported to increase with exercise training. In the same sense, NON had the lowest intrinsic endothelial function among four inbred strains; therefore, exercise training would be expected to improve endothelial function more in NON than other strains (higher trainability for endothelial function).
Our findings of variation in vascular responses to exercise training across strains are consistent with variable responses to training observed in humans and other animals. Improvement in endothelial function with exercise training has been reported previously [2345]. Conversely, other studies have reported that endothelial function is not improved by exercise training in young healthy humans and animals [2945464748]. For example, Batacan et al. [48] reported no improvement in EDR in aortic and mesenteric arterial rings from rats completing 12 weeks of low (40% of max) or high (> 90% of max) intensity exercise training compared to sedentary rats. Haram et al. also showed no differences in maximal responses to ACh in aortas from aerobic interval- or moderately-trained rats and sedentary controls [29]. Thus, the endothelial responses to exercise training are variable and likely related to baseline endothelial function and the exercise volume (e.g., intensity). The results of the current study also revealed that genetic background is another potential determinant of the vascular responses to exercise training.
In many studies, HIT has been proposed as an effective alternative to the traditional aerobic training, inducing similar or even superior cardiovascular adaptations [3192029303249]. In the present study, EDR was improved similarly in MOD and HIT of NON, suggesting that HIT could be a time-efficient way to improve endothelial function. However, very interestingly, SJL HIT had impaired EDR compared to SED (Fig. 3C). Previous studies reported that intense exercise caused a decrease in eNOS expression and NO level in hearts of young rats [26] and induced platelet aggregation in young healthy humans, which may augment the risk of vascular thrombosis [50]. Bergholm et al. [25] found that vigorous physical training (70%–80% of VO2max) for 12 weeks impaired endothelium-dependent vasodilation and decreased antioxidant concentrations in young individuals. Also, high intensity exercise training-induced an increase in plasma concentration of 8-hydroxy-2′-deoxyguanosine, a marker of oxidative stress to DNA, without a beneficial effect on endothelial function [27]. Although nitrotyrosine was measured in skeletal muscle, the higher nitrotyrosine levels in HIT could reflect a higher systemic oxidative stress level in this group, explaining the impaired endothelial function in SJL HIT (Fig. 7). Our finding supports the previously proposed concept that a threshold of training intensity exists beyond which ROS generation overrides the scavenging capabilities of antioxidant systems in the vasculature [42]. Therefore, comparing the different effects of HIT on endothelial function between NON and SJL in the present study, the beneficial threshold of training intensity would be determined by genetic background. Alternatively, the response pattern observed in SJL HIT is similar to that observed in mice completing overtraining protocols [51]. However, we did not measure other biological markers of overtraining in these mice, therefore a future study would be required to characterize the responses in SJL HIT as overtraining.
The number of differentially expressed genes was proportional to the difference in EDR between SED and exercise-trained groups (Tables 3 and 4). Furthermore, the number of genes differentially expressed in HIT was greater than MOD in all three strains and only a few genes overlapped between MOD and HIT. This observation is opposite to a previous study that moderate intensity endurance training produced greater transcriptional effects on isolated aortic endothelial cells from rats than interval sprint training [52]. However, those authors also reported that interval sprint training had a greater effect on gene expression in skeletal muscle feed arteries than endurance exercise training, implying that transcriptional responses to exercise training could be dependent on species as well as vessel characteristics and training intensity.
Differences in training intensity can influence not only the number of genes differentially expressed, but also the signaling pathways in which those genes are involved. A previous study suggested that different intensity training programs exert shear stress on the vessel walls differently during exercise and this yields differences in molecular responses [31]. Similarly, Padilla et al. [52] reported no overlap for gene networks influenced by endurance and interval sprint training for skeletal muscle feed arteries and rat aortic endothelial cells. Thus, our findings provide further evidence for the intensity-specific pattern of training-induced transcriptional activation in mouse aortas.
The biological pathways and molecular/cellular functions overrepresented with genes differentially expressed in aorta varied by exercise intensity (Tables 5 and 6). A gene set differentially expressed in SJL HIT was anticipated to increase ROS production. Additionally, the top disease identified by IPA analysis for the gene set was ‘aortic aneurysm’ (data not shown, p = 8.65E-03). These findings are consistent with the impaired endothelial function observed in SJL HIT and are in line with a previous study that endothelial genes altered by exercise training in swine vessels were associated with leukocyte migration and/or chemotaxis biological processes [53]. In contrast, a gene cluster differentially expressed in NON HIT was predicted to decrease adhesion and migration of endothelial cells (Table 6). The prediction fits with the concept that exercise training is associated with athero-protective changes in the endothelium including decreased adhesiveness for inflammatory molecules [54].
Prolonged exercise training can induce vascular remodeling, resulting in larger blood vessels [555]. In NON MOD, differentially expressed genes were connected to molecular/cellular functions of increased endothelial cell development and proliferation (Table 6). With the notion that outward vascular remodeling of blood vessels in response to exercise training can lead to decreased acute responses to endothelium-dependent vasodilators and increased blood flow [59], NON MOD might elicit a transcriptional response leading to vessel remodeling after a more prolonged training protocol.
In the present study, intrinsic CS activity varied across strains (Fig. 1A), suggesting a genetic contribution to intrinsic CS activity in mouse skeletal muscle. This finding is in agreement with Ratkevicius et al. in that there is a difference in intrinsic CS activity among inbred mouse strains [56]. Those authors identified a putative single nucleotide polymorphism (rs29358506) that contributed to the difference in intrinsic CS activity across 6 inbred mouse strains. We could not support their findings because all four strains utilized in the present study have the same genotype for this polymorphism. For SDH activity, we found no difference among four inbred strains (Fig. 1B) and thus do not have evidence of a genetic contribution to intrinsic SDH activity.
Although we demonstrated that exercise training has non-uniform effects on endothelial function depending on the interaction between genetic background and training intensity, our study has some limitations. First, the four strains utilized in the current study are genetically diverse [33]; however, the genetic diversity in these four strains is limited relative to humans. Therefore, the phenotypic variation in our study might not be directly translatable to individual variation observed in humans. Nevertheless, the strain differences in endothelial function and transcriptional activation of endothelial genes in this limited number of strains supports the use of more genetically diverse mouse populations (e.g., Diversity Outbred mice) to investigate this variation. Second, other exercise training components were not considered in the current study. For instance, training duration is also an important factor critically influencing training effects on vascular function [42]. It might be speculated that genetic factors possibly interact with training duration as well for the effects of exercise training on endothelial function. Third, only thoracic aortas were utilized in the current study. Many studies have indicated that endothelial responses to exercise training are heterogeneous across the vessel sizes and locations [5], raising the possibility that genetic contribution to endothelial responses to exercise training could be vessel-specific. In addition, the sex difference has been suggested as a factor determining endothelial responses to exercise training [5758], implying the potential difference in genetic regulation of endothelial responses to exercise training between sexes. Accordingly, proposed vessel- and sex-specific differences in genetic contribution to endothelial responses to exercise training represent an intriguing area for future research.
In summary, we demonstrated that intrinsic vascular function in mouse aortas, mitochondria oxidative enzyme activity and nitrotyrosine abundance in skeletal muscle are variable across inbred mouse strains, suggesting a genetic influence. Most importantly, we report a novel finding that exercise training has non-uniform effects on endothelial function and transcriptional activation of endothelial genes depending on the interaction between genetic background and training intensity. These results indicate that the optimal intensity of exercise training to improve endothelial function could vary by individuals. Results from skeletal muscle oxidative enzyme activity, the abundance of oxidative stress and pathway analysis for gene expression profiles further support the intricate interaction between genetic background and training intensity on responses to exercise training. The present study, for the first time, provides evidence of an interactive effect between genetic background and training intensity on exerciseinduced vascular adaptation. Further studies incorporating larger scale expression profiles, for example, RNAseq, microarray or full sequencing, on different genetic backgrounds might help expand our knowledge of the mechanism regulating endothelial responses to exercise training.
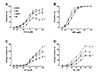









 XML Download
XML Download