

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Ezrin, radixin, and moesin proteins, collectively called as ERM, are a group of proteins with high degree of homology among themselves [1], and play important roles in cell-shape determination and structural stability by linking actin cytoskeleton to plasma membrane [23]. When phosphorylated at a threonine residue at C-terminal regulatory domain, they become activated and bring F-actin close to plasma membrane, where they act as important mediators of cellular signaling pathways between outer and inner cellular compartments [14]. Thus, ERM proteins reside in a position that contribute to structural plasticity and changes of molecular signaling pathways linked to cytoskeletons [356].
The nucleus accumbens (NAcc) is the brain site where drugs of abuse produce their rewarding effects [789]. The molecular changes in signaling pathways in this site are known to be critical for the expression of addictive behaviors [10]. Previously, we have found that ERM phosphorylation in this site is dose-dependently reduced by acute injection of cocaine [11], and especially in the core, a sub-region of the NAcc, by amphetamine (AMPH) [12]. Considering that ERM proteins are critical for the cellular structure, these findings suggest that ERM proteins may play a role in structural remodeling by taking a part in a series of psychomotor stimulant-induced cellular events in the NAcc.
While there is no information at all about how ERM proteins are regulated in the NAcc, we have previously shown that AMPH-induced decrease of ERM phosphorylation in the NAcc core, but not in the shell, are reversed by lithium [12]. Interestingly, it is shown that lithium activates protein kinase B (Akt) and subsequently inhibits glycogen synthase kinase 3β (GSK3β) by increasing their phosphorylation [131415]. Put together, it is plausible to make a hypothesis that a signaling pathway mediated by Akt-GSK3β may also regulate ERM phosphorylation in the NAcc core. Supporting this possibility, it has been shown, although in neuronal and epidermal cell cultures, that ERM phosphorylation is under the regulation of Akt and its upstream effector, phosphatidylinositol 3-kinase (PI3 kinase) [1617]. More recently, it has been also shown in murine auditory epithelial progenitor cells that Akt activation increases ERM phosphorylation levels [18]. Thus, we examined in the present study whether direct manipulation of Akt-GSK3β signaling in the NAcc core may influence ERM phosphorylation in this site.
METHODS
Subjects
Male Sprague–Dawley rats weighing 220–250 g (equivalent to 6 weeks olds) on arrival were obtained from Orient Bio Inc. (Seongnam-si, Korea). They were housed 3 per cage in a 12 h light/dark cycle room (lights out at 8:00 PM), and all experiments were conducted during the day time. The rats had access to food and water ad libitum at all times. All animal use procedures were conducted according to an approved Institutional Animal Care and Use Committee protocol (approval number: 09-140).
Drugs and peptide
LY294002 hydrochloride (Sigma-Aldrich, St. Louis, MO, USA), PI3 kinase inhibitor, was dissolved in 100% DMSO and small aliquots were stored at −80℃. Immediately before use, frozen aliquots were diluted to final working concentrations of 0.8 or 8.0 µg/µl in 80% DMSO. S9 peptide, which consists of 21 amino acids (a.a.) including a small peptide (11 a.a.), commonly referred to as protein transduction domain [19], and a portion (10 a.a.) of the N-terminus sequence of GSK3β (GRPRTTSFAE) known as the substrate site for Akt and thereby competes with GSK3β against its phosphorylation [2021], was artificially synthesized and kindly provided by Professor Soo Young Lee at the Center for Cell Signaling and Drug Discovery Research, Ewha Womans University (Seoul, Korea). It was dissolved to final working concentrations of 1.0 or 10.0 µg/µl in saline.
Surgical procedures
Rats were anesthetized with intraperitoneal ketamine (100 mg/kg) and xylazine (6 mg/kg), placed in a stereotaxic instrument with the incisor bar at 5.0 mm above the interaural line and implanted with chronic bilateral guide cannulas (22 gauge tubing size with 0.71 mm in outer and 0.39 mm in internal diameters; Plastics One, Roanoke, VA, USA) aimed at the NAcc core (A/P, +3.4; L, ±1.5; D/V, −7.5 mm from bregma and skull) [22]. Cannulas were angled at 10° to the vertical, positioned 1 mm above the final injection site, and secured with dental acrylic cement anchored to stainless steel screws fixed to the skull. After surgery, 28 gauge (tubing size with 0.36 mm in outer and 0.18 mm in internal diameters) obturators were placed in the guide cannulas, and rats were returned to their home cages for a 7-day recovery period.
Design and procedures
Upon arrival, all rats passed a week-long adaptation period to the new housing environment. Then, bilateral guide cannula surgery was followed. About 1 week after surgery, three different groups of rats were bilaterally microinjected into the NAcc core with vehicle (80% DMSO) or LY294002 (0.4 or 4.0 µg/0.5 µl/side) (experiment 1), while a separate set of three different groups of rats were bilaterally microinjected into the NAcc core with vehicle (saline) or S9 peptide (0.5 or 5.0 µg/0.5 µl/side) (experiment 2). Bilateral intracranial microinjections into the NAcc core were made in the freely moving rat. Injection cannulas (28 gauge) connected to 1 µl syringes (Hamilton, Reno, NV, USA) via PE-20 tubing were inserted to a depth 1 mm below the guide cannula tips. Injections were made in a volume of 0.5 µl per side over 30 sec. After 1 min, the injection cannulas were withdrawn and the obturators were replaced. Then, they were placed back to cage and let them rest for 60 min, decapitated and the brain tissues (core only) were prepared for western blot analysis.
Brain tissue preparation
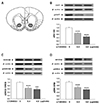
Animals were decapitated by guillotine 60 min after vehicle, drug, or peptide microinjections. Brains were rapidly removed and coronal sections (1.0 mm thick extending 1.60–2.60 mm from bregma) were obtained with an ice-cold brain slicer (Model SA-2160, Roboz Surgical Instrument Co., Gaithersburg, MD, USA). The NAcc core tissues were obtained in the circular punch with 1.2 mm diameter on an ice-cold plate (Fig. 1A), immediately frozen on dry ice and stored at −80℃. They were prepared bilaterally and pooled for each individual animal's protein isolation.
Western blotting
Tissues were homogenized in lysis buffer containing 0.32 M sucrose, 2 mM EDTA, 1% SDS, 10 µg/ml aprotinin, 10 µg/ml leupeptin, 1mM phenylmethylsulfonyl fluoride, 10 mM sodium fluoride, and 1 mM sodium orthovanadate. The concentration of protein was determined by using Pierce Coomassie Protein Assay Kit (Thermo Scientific Inc., Rockford, IL, USA). Samples were then boiled for 10 min and subjected to SDS-polyacrylamide gel electrophoresis. Proteins were separated and transferred electrophoretically to nitrocellulose membranes (Bio-Rad, Hercules, CA, USA), which were then blocked with 5% bovine serum albumin (BSA) in PBS-T buffer (10 mM phosphate-buffered saline plus 0.05% Tween-20). Antibodies used to probe the blots were as following: total Akt (1:10,000), phosphor-Akt (1:500), total GSK3β (1:4,000), phosphor-GSK3β (specific to detect phosphorylated GSK3β at serine 9, 1:2,000), total ERM (1:1,000), phosphor-ERM (specific to detect phosphorylated ezrin-radixin-moesin at threonine 567, 564 or 558, respectively; 1:500), purchased from Cell Signaling (Beverly, MA, USA) and diluted in PBS-T with 5% BSA; β-actin (1:10,000), purchased from Abcam (Cambridge, UK) and diluted in PBS-T with 5% skim milk. Two separate gels were used to detect total and phosphorylated proteins, respectively. Primary antibodies were detected with peroxidase-conjugated secondary antibodies, anti-rabbit IgG (1:2,000; KOMA Biotech, Seoul, Korea) diluted in PBS-T with 5% skim milk, followed by enhanced chemiluminescence reagents (Amersham Biosciences, Arlington Heights, IL, USA) and exposure to X-ray film. Band intensities were quantified based on densitometric values using Fujifilm Science Lab 97 Image Gauge software (version 2.54) (Fujifilm, Tokyo, Japan).
Statistical analyses
Statistical analyses were performed using the Sigma Plot version 12.0 (Systat Software, San Jose, CA, USA). The data were analyzed with one-way ANOVA, followed by post-hoc Bonferroni comparisons. Differences between experimental conditions were considered statistically significant when p < 0.05.
RESULTS
Microinjection of LY294002 into the NAcc core decreases ERK phosphorylation levels in this site
In order to examine our hypothesis that ERM phosphorylation is under the regulation of Akt and GSK3β in the NAcc core, LY294002 was bilaterally microinjected into this site and phosphorylation levels for each molecules were measured. As expected, microinjection of LY294002 into the NAcc core decreased phosphorylation levels of Akt, and subsequently of GSK3β, in this site (Fig. 1B, C). The one-way ANOVA conducted on these results revealed significant effects of treatment (F2,15 = 57.49, p < 0.001, for Akt; F2,15 = 13.53, p < 0.001, for GSK3β, respectively). Further, microinjection of LY294002 into the NAcc core also decreased the phosphorylation levels of ERM in this site (Fig. 1D). The one-way ANOVA conducted on these results showed significant effects between different doses of drugs (F2,15 = 25.05, p < 0.001).
Microinjection of S9 peptide into the NAcc core decreases ERK phosphorylation levels in this site
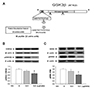
In order to confirm whether reduction of GSK3β phosphorylation in the NAcc core actually contributed to the decrease of ERM phosphorylation in this site, we microinjected S9 (see Fig. 2A) bilaterally into the NAcc core with a separate group of rats and measured the phosphorylation levels of GSK3β and ERM. The one-way ANOVA conducted on these results revealed significant effects of treatment (F2,14 = 4.59, p < 0.05, for GSK3β; F2,14 = 4.39, p < 0.05, for ERM, respectively). Consistent with our previous findings [23], we found that microinjection of S9 into the NAcc core dose-dependently decreased GSK3β phosphorylation in this site (p < 0.05 by post-hoc Bonferroni comparisons; higher dose of S9 versus saline) (Fig. 2B). Interestingly, we also found that microinjection of S9 dose-dependently decreased ERM phosphorylation in this site (p < 0.05 by post-hoc Bonferroni comparisons; higher dose of ERM versus saline) (Fig. 2C).
DISCUSSION
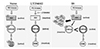
The present results demonstrate that ERM phosphorylation in the NAcc core is decreased by microinjections of S9 peptide, a proposed GSK3β activator, as well as LY294002, PI3 kinase inhibitor, in this site. This is the first direct demonstration, to our knowledge, to indicate that ERM is under the regulation of Akt-GSK3β signaling in the NAcc core.
There has been no study at all in the literature about what molecular signaling pathways regulate phosphorylation levels of ERM proteins in the NAcc. However, it has been previously shown in our lab that ERM phosphorylation is decreased by acute AMPH in the NAcc core, and interestingly it is recovered back to saline control level when lithium is co-present [12]. Because lithium, among multiple pathways that it affects [24], is known to increase phosphorylation of Akt and GSK3β [1314], these results plausibly suggest that Akt-GSK3β signaling pathway may position to regulate ERM phosphorylation in the NAcc core. As we hypothesized, we showed in the present study that direct (by reduction of Akt phosphorylation) or indirect (by competition with Akt against GSK3β phosphorylation) reduction of GSK3β phosphorylation levels in the NAcc core actually produces the decrease of ERM phosphorylation in this site (see Fig. 3). Although it has been previously shown in neuronal and epidermal cell cultures, and more recently in PC12 cells, that PI3 kinase-Akt signaling pathway, which usually positioned as upstream effectors of GSK3β, regulate ERM phosphorylation levels [161725], it is first time to directly show in the NAcc that this pathway also regulates ERM signaling in this region.
It is interesting to compare our present data with a recent report by Jin et al. (2018) [25]. They found that an Akt inhibitor specifically decreased radixin protein levels, but not ezrin and moesin, in PC12 cells. Because we didn't discriminate radixin out of other ERM family proteins, we are not sure whether it would be the case in the NAcc. Nevertheless, it is worth to mention that there is an obvious difference in that they measured protein levels in PC12 cells (in vitro, homogenous), while we did it in the NAcc tissues (in vivo, heterogenous). Interestingly, we previously reported [26] that AMPH rather increases a ratio of phosphorylated to total ERM in this cell, which is opposite of what we observed in the NAcc tissue [12]. Therefore, we think that there might be different regulatory mechanism working depending on the type of cell or brain area that we are looking.
Next, as a down-stream effector of Akt, it has been shown that phosphorylation levels of GSK3β are decreased in the NAcc core by acute cocaine [23]. In parallel, we have found that ERM phosphorylation is also decreased in this site by either acute cocaine [11] or acute AMPH [12]. Put together with these results, the present findings suggest that the effect of psychomotor stimulants on the reduction of ERM phosphorylation in the NAcc core is possibly through a phasic regulation of Akt-GSK3β signaling pathway.
As activation of ERM proteins requires the phosphorylation at the C-terminal region [13], the decrease of phosphorylation levels for these proteins in the NAcc core by psychomotor stimulants as shown in our previous reports [1112] indicates that there might be a chance for a structural change to occur in this region. Supporting this possibility, it was actually shown that dendritic spine density in the NAcc was increased by acute cocaine after 6 h [27]. Although the detailed mechanism about what molecular pathways are involved in this process is not known, it may be possible that the reduction of the influence of Akt-GSK3β signaling pathway in response to acute psychomotor stimulants may result in decrease of ERM phosphorylation, which then induces a series of cellular events leading to collapse and remodeling of actin cytoskeletons to subsequently produce dendritic spine changes. Even it may be transient, such a change may become a form of metaplasticity [28], and thereby contribute to more endurable structural plasticity in the long run, eventually leading to behavioral changes manifested as addictive behaviors when repeatedly exposed to drugs of abuse [293031]. These possibilities remain open to be explored in the future.
In conclusion, the present findings showed that ERM phosphorylation in the NAcc core is under the regulation of Akt-GSK3β signaling pathways, and suggest that they may be positioned as upstream effectors to ERM. As it is not mostly explored yet what ERM proteins function in the brain, especially in the NAcc, our present findings will be a small step toward our understanding for ERM proteins in terms of revealing their signaling pathways in this region. Further, considering their role in structure formation and maintenance, revealing ERM-related signaling pathway in the NAcc, an important region mediating addictive behaviors, as shown in the present findings, has a significant implication to understand structural plasticity and relevant signaling in drug addiction.
 XML Download
XML Download