

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Ion channels in arterial smooth muscle regulate the membrane potential, Ca2+ permeability, and vascular tone, and maintain homeostasis [1]. There are several types of K+ channels in arterial smooth muscle cells (SMCs). Voltage-gated K+ channels (Kv) are activated by membrane depolarization to induce membrane repolarization and prevent excessive Ca2+ influx through L-type Ca2+ channels [23]. Inward rectifier K+ channels (Kir) contribute to the establishment of a hyperpolarized membrane potential and mediate endothelium-dependent hyperpolarization and the active hyperemia response in various vessels, including the coronary artery (CA) [45]. Big-conductance Ca2+-activated K+ channels (BKCa) are abundantly expressed in SMCs and are activated in concert by membrane depolarization and increased concentrations of cytosolic Ca2+ ([Ca2+]c) to provide a negative feedback regulatory component in the arterial smooth muscle [6].
Compared with the current knowledge of K+ channels in arterial smooth muscle, our understanding of K+ channels in the CA, especially regarding the branches perfusing different areas of the heart, is limited. As major trunks of the CA originating from the initial segment of the aorta, the left coronary arteries (LCAs)—which are divided into left anterior descending (LAD) and left circumflex coronary (LCX) arteries—are located between the pulmonary artery and the left atrial appendage and flow to the left ventricular (LV) walls and partly, to the septum. The right coronary arteries (RCAs) are located on the right side of the heart and perfuse the right ventricular (RV) walls and the septal area. Septal coronary arteries (SCAs) originate from both LCAs and RCAs [7].
Coronary circulation provides oxygen and nutrients to the cardiomyocytes, and its impairment causes major cardiac diseases, such as ischemic heart disease [8]. Several studies have demonstrated that impaired coronary circulation is also related to right heart diseases associated with pulmonary arterial hypertension (PAH) and RV hypertrophy (RVH) [91011]. PAH is a lethal vascular disease characterized by high pulmonary arterial pressure (> 25 mmHg) and progressive remodeling of the pulmonary vasculature [12]. The increased afterload to the RV causes RVH in PAH patients and can inhibit adequate CA blood flow to the RV, thereby increasing the risk of ischemia and RV failure. Various animal models have been used to identify the underlying mechanisms associated with PAH [13]. Among these, the monocrotaline (MCT)-induced PAH model involving single injection of MCT in rats has been widely used [14], with numerous studies reporting the functional roles of pulmonary arterial smooth muscle and endothelial dysfunction in MCT-induced PAH (MCT-PAH) [151617].
Interestingly, a recent study reported CA remodeling, including thickening of the vascular wall, in patients with PAH as well as in MCT-PAH models [18]. Given the plausible elevated risk of ischemic damage to the RV, branch-specific changes in K+ channel currents in CA smooth muscle would be an intriguing research topic; however, most studies on arterial smooth muscle in PAH have focused on pulmonary arteries [151617].
Regarding the comparison of K+ channel activities between RCAs and LCAs, a previous study demonstrated higher levels of Kv1.2-type current showing slow inactivation in RCA SMCs (RCSMCs) than that in LCA SMCs (LCSMCs) of healthy Wistar rats [19]. In contrast, another study reported that non-inactivating Kv7-type current was increased in LCSMCs than that in RCSMCs of healthy rats [1920]. Moreover, a previous study using a PAH disease model of chronic hypoxia showed that Kv1.2- and Kv1.5-type currents were decreased in RCAs than those in LCAs [21].
In the present study, we elucidated the changes in K+ channel currents in CA myocytes isolated from MCT-PAH model rats, with special focus on branch-specific comparisons. We initially compared the densities of three major types of K+ channel current (IKv, IKir, IBKCa) between LCSMCs, RCSMCs, and SMCs from the septal branch from the main trunk of the right CA (SCSMCs).
METHODS
Animals
All experimental procedures were performed with the approval of the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (IACUC approval no: 170626-3-1). Male Sprague–Dawley rats (230–280 g) were used for all experiments. MCT-induced PAH was developed by a single intraperitoneal (i.p.) injection of MCT (60 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) into 7-to-8-week-old rats; control rats were injected with an equal amount of saline. After 3 weeks, MCT-injected rats (hereafter referred to as MCT-PAH rats) were euthanized. All rats were anaesthetized by i.p. injection of a ketamine and xylazine mixture (ketamine 90 mg/kg and xylazine 10 mg/kg).
RVH histology
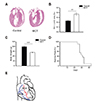
For histological analysis, hearts were fixed in 4% paraformaldehyde overnight, and paraffin-embedded tissue sections were cut (5 µm thickness) and stained with hematoxylin and eosin (H&E). Digital images of stained tissues were obtained at a magnification of 200× using an Olympus BX51 (Olympus, Tokyo, Japan) and Photoshop 7.0 software (Adobe, San Jose, CA, USA) (Fig. 1A). The RVH index was calculated as a ratio of the RV weight versus the composite weight of the LV and septum (S) as follows: RVH index = RV/(LV+S) (Fig. 1B).
Preparation of vessels and single myocyte isolation
CAs were dissected under a surgical microscope (SMZ645; Nikon, Tokyo, Japan). The LCAs used in this study included first and second branches of the LAD and LCX arteries located on the surface of the heart. In addition, RCA branches were dissected from the surface of the RV, and SCAs were dissected from the surface of the internal cavity of the RV (Fig. 1E). After dissection of the arteries, vessels were incubated in the first digestion medium (nominal Ca2+-free normal Tyrode (NT) solution containing 1 mg/ml papain; Sigma-Aldrich) for 17 min, followed by incubation in the second digestion medium (nominal Ca2+-free NT solution containing 3 mg/ml collagenase; Wako Pure Chemical, Osaka, Japan) for 13 min. Both digestion media contained bovine serum albumin (1 mg/ml; Calbiochem, Billerica, MA, USA) and dithiothreitol (1 mg/ml; Sigma-Aldrich).
Electrophysiology
Whole-cell patch-clamp experiments were performed using a patch-clamp amplifier (Axopatch-200A; Axon Instruments, Foster City, CA, USA). We used pCLAMP software (v.10.4) and Digidata-1440A (Axon Instruments) to acquire data and to apply the command pulse. Current recordings were made at room temperature (23℃–25℃), using glass microelectrodes with a resistance of approximately 2.5–3.0 MΩ. Membrane capacitance of each cell was measured and was used to normalize the current amplitude (i.e., current density [pA/pF]).
Experimental solutions and chemicals
The pipette solution for recording IKv and IKir contained 120 mM K-aspartate, 20 mM KCl, 10 mM HEPES, 1 mM MgCl2, 3 mM MgATP, and 5 mM EGTA (adjusted to pH 7.2 with KOH). The NT external solution comprised 140 mM NaCl, 5.4 mM KCl, 10 mM HEPES, 10 mM glucose, 1 mM MgCl2, and 1.8 mM CaCl2 (adjusted to pH 7.4 with NaOH). For IKir, symmetric high-K+ 140 mM KCl bath solution (KCl solution) was used (140 mM KCl, 10 mM HEPES, 10 mM glucose, 1.8 mM CaCl2, and 1 mM MgCl2 [adjusted to pH 7.4 with KOH]). After confirming the steady-state current under symmetrical high-K+ conditions, 100 µM BaCl2 was administered to block the inward K+ current component in order to selectively calculate the density of IKir. For the IBKCa channel recording, the pipette solution contained 120 mM K aspartate, 20 mM KCl, 10 mM HEPES, 3 mM MgATP, 10 mM EGTA, 1 mM MgCl2, and 8.6 mM CaCl2 (pH 7.2), with the free Ca2+ concentration fixed at 1 µM.
RESULTS
Compared with control rats, MCT rats showed a thickened RV wall (Fig. 1A), as well as a significantly increased RVH index [RV/(LV+S)] (control rats: 0.2 ± 0.01, n = 7; MCT rats: 0.4 ± 0.03, n = 6; p < 0.01) (Fig. 1B). At 3 weeks after MCT injection, the body weights (g) of MCT-injected rats were lower than those of agematched controls (control rats: 119.2 ± 4.02 g, n = 36; MCT rats: 83.7 ± 2.71, n = 37; p < 0.001) (Fig. 1C). At 30 days after injection, < 10% of MCT rats survived (Fig. 1D).
Smaller IKv in SCSMCs
In the whole-cell patch-clamp configuration study, IKv was activated by membrane depolarization from a −80 mV holding voltage with 5 mM EGTA-containing KCl pipette solution. Step-like depolarization ranging from −50 mV to 40 mV induced outward currents showing slow inactivation during a 2.5-s pulse period (Fig. 2). Peak outward currents normalized to the membrane capacitance (i.e., pA/pF) were determined in order to compare the level of Kv activity. Although a buffer with high-concentration of Ca2+ (5 mM EGTA) was included in the pipette solution, we also examined the effect of paxilline, a highly selective and potent blocker of BKCa, in order to exclude IBKCa activation by strong depolarization. Paxilline application did not affect the amplitudes of the peak outward currents, indicating that the recordings obtained using the 5 mM EGTA pipette solution were not contaminated by IBKCa (Fig. 2G).
The IKv densities were similar in control LCSMCs (12.7 ± 1.45 pA/pF at +40 mV, n = 27) and control SCSMCs (12.3 ± 1.08 pA/pF at +40 mV, n = 15), but higher in control RCSMCs (19.8 ± 2.44 pA/pF at +40 mV, n = 17) (Fig. 2H). Comparison of control and MCT-PAH rats revealed that the IKv densities in LCSMCs and RCSMCs did not differ (Fig. 2A, B, E, F), whereas they were lower in SCSMCs from MCT-PAH rats than in those from control rats (12.3 ± 1.08 pA/pF vs. 8.8 ± 0.95 pA/pF at +40 mV, n = 15) (Fig. 2C, D). Additionally, the I/V curves showed that the current densities in MCT-PAH SCSMCs were significantly decreased at 20 mV, 30 mV, and 40 mV (p < 0.05) (Fig. 2D), and that the current densities in control RCSMCs were significantly greater than those in control LCSMCs (p < 0.05) and SCSMCs (p < 0.01).
Voltage-dependent inactivation of IKv
The raw traces of IKv revealed slow inactivation during the depolarizing pulses (Fig. 2A, C, E). Because the resting membrane potential of vascular smooth muscle usually exists at relatively depolarized levels (i.e., −40 to −60 mV), the steady-state level of partially inactivated IKv would represent the effective outward current. Therefore, we analyzed the voltage dependence of inactivation using a double-pulse protocol. Different levels of long prepulses (10 s) were applied, immediately followed by a +40-mV test pulse for 30 ms (Fig. 3A). The normalized amplitudes at the test pulse were plotted against the various depolarizing pre-pulses, which reflected the voltage-dependent inactivation. The voltage dependences illustrated by the inactivation curves were obtained by fitting the data to the Boltzmann equation. The half-inactivation voltage (V1/2) of control SCSMCs (−40.1 ± 1.58 mV) was more negative than that of control LCSMCs (−31.0 ± 0.74 mV) and RCSMCs (−29.3 ± 1.51 mV) (Fig. 3B). Interestingly, the V1/2 of MCT-PAH SCSMCs was shifted slightly toward the right (−35.0 ± 0.83 mV), and V1/2 was not significantly different between MCT-PAH LCSMCs (−30.6 ± 0.86 mV) and MCT-PAH RCSMCs (−30.3 ± 1.62 mV) (Fig. 3C).
Next, we analyzed the voltage-dependent activation property using the Boltzmann equation after plotting the normalized conductance (G/Gmax) of outward currents against the test voltages. The voltage-dependent activation tended to be similar between control and MCT-PAH rats. The V1/2 of both control LCSMCs (−6.1 ± 1.61 mV) and MCT LCSMCs (−5.5 ± 1.33 mV) was shifted toward the left, whereas that of SMCs from other arteries was not (Fig. 3D, E). Control RCSMCs (5.8 ± 0.79 mV) and control SCSMCs (6.6 ± 2.12 mV) had similar current densities, as did the MCT RCSMCs (4.6 ± 2.01 mV) and MCT SCSMCs (3.1 ± 2.27 mV).
IKir in CA myocytes from control and PAH-MCT rats
For selective recording of IKir, the bath solution was changed from an NT to a KCl solution, and the membrane voltage was maintained at 0 mV. We applied a ramp-like hyperpolarizing pulse from 30 to −140 mV, and the brief I/V curve obtained by the ramp pulse showed an inwardly rectifying current, which was abolished by applying 100 µM Ba2+, a Kir blocker (Fig. 4A). The Ba2+-sensitive current reflecting IKir was obtained by digital subtraction and compared between the three branches (Fig. 4B). The IKir densities at −140 mV were significantly smaller in control SCSMCs (−11.9 ± 2.46 pA/pF, n = 13, p < 0.05) than that in control LCSMCs (−18.0 ± 1.76 pA/pF, n = 17) and control RCSMCs (−17.1 ± 2.47 pA/pF, n = 16) (Fig. 4C). Interestingly, the IKir densities of MCT-PAH SCSMCs (−5.0 ± 0.59 pA/pF, n = 21) were even smaller than those of control SCSMCs (−11.9 ± 2.46 pA/pF, n = 13; p < 0.001) (Fig. 4D). In contrast, the IKir densities of RCSMCs and LCSMCs did not differ between the control and MCT-PAH (Fig. 4D).
IBK of CA myocytes from control and MCT-PAH
To record IBKCa, we used a KCl pipette solution with 1 mM of free Ca2+ buffered with 10 mM EGTA. Because BKCa is not inactivated at the depolarized membrane potential, we applied a ramp-like hyperpolarizing pulse from 50 mV to −100 mV at the depolarized holding voltage (−10 mV) in order to induce IKv inactivation. The outwardly rectifying current was completely abolished by addition of 1 µM of paxilline, indicating selective recording of IBKCa in CA myocytes (Fig. 5A). IBKCa density did not differ between myocytes of the three CA branches in controls (Fig. 5B), and no significant difference in IBKCa was observed between control and MCT-PAH CA myocytes (Fig. 5C–E).
DISCUSSION
We investigated the functional remodeling of the three types of K+ channel currents in SMCs obtained from the three branches of the CA in MCT-PAH and control rats. Although we initially hypothesized that the measured parameters might differ to a greater degree in RCSMCs than that in LCSMCs and SCSMCs, we mainly observed intriguing differences in SCSMCs.
Different levels of IKv among CA branches from control rats and changes associated with PAH
Electrophysiological analyses of control rats demonstrated branch-specific differences in K+ currents, including higher IKv in RCSMCs and lower IKir in SCSMCs (Figs. 2H and 4C). Similarly, a previous comparative study of IKv between RCSMCs and LCSMCs reported a higher peak amplitude in RCSMCs than that in LCSMCs [19], although SCSMCs were not investigated in this study. Another study revealed that the Kv7-type current of IKv was larger in LCSMCs than that in RCSMCs, but total Kv currents were not compared between LCA and RCA [20], which makes it difficult to compare these findings with our results. We attempted to measure and compare Kv7-sensitive currents using linopirdine, a Kv7 inhibitor; however, linopiridine-sensitive currents were too small to distinguish the differences between LCA and RCA (data not shown). A previous study that compared Kv7 currents in the cerebral artery and CA reported that responses of Kv7 currents in CA SMCs to linopirdine were blunted when compared to those in cerebral artery SMCs, and Kv7 component in total K+ conductance was relatively small [22]. Our results partially agree with those reported by Gautier et al. [19], in that the peak amplitude of IKv in control RCSMCs was larger than that in the other branches (Fig. 2H). However, because the difference in IKv was not significant at the physiological range of negative membrane voltages, the physiological implications of the higher IKv density in vivo remain to be explored. Rather than the peak amplitude, the difference in steady-state inactivation at the negative membrane voltage requires further attention. The IKv in SCSMCs showed more sensitive voltage-dependent inactivation (i.e., a more negative V1/2), suggesting a relatively smaller steady-state conductance in SCSMCs than that in the other branches. Furthermore, the IKir density in SCSMCs was smaller than that in RCSMCs and LCSMCs.
Previous comparisons of IKv between RCAs and LCAs were conducted in rat models of hyperglycemia and chronic hypoxia [2021]. Hyperglycemic rats showed a decrease in Kv7 current density, which was more prominent in LCSMCs than in RCSMCs [20], whereas in a chronic hypoxia state that led to PAH, Kv current densities were decreased in RCSMCs, but increased in LCSMCs [21]. Notably, these studies did not examine the IKv in SCSMCs. In the present study, using CA myocytes from MCT-PAH rats, we observed no significant changes in the IKv in LCSMCs and RCSMCs, whereas that in SCSMCs consistently decreased. However, we noted a slight shift to the rightward direction for the inactivation curve for SCSMCs (Fig. 3B, C), which might have partly compensated for the decrease in the peak current density of IKv. The discrepant results might be explained by the different types of disease models.
Different levels of IKir among CA branches from control rats and changes associated with PAH
Kir in arterial myocytes are generally more abundant in vessels with a smaller diameter, and thereby contribute to the resting membrane potential and basal tone of the arteries [23]. Although the functional expression of Kir in CA myocytes has been reported in pigs [23], rabbits [24], and rats [25], differences in IKir among the major branches of CAs have not been studied. Furthermore, to the best of our knowledge, decreased IKir in CA myocytes from PAH animal models has never been investigated.
Kir activity is facilitated by extracellular K+ activity ([K+]e) and represents the underlying mechanism of membrane hyperpolarization under moderate increases in [K+]e. Moreover, the hyperpolarization and subsequent relaxation of Kir-expressing arteries underlie [K+]e-induced vasorelaxation, which is critical for matching the regional blood flow to electrically excitable tissues, such as myocardium. In this respect, the lower IKir observed in SCSMCs and its attenuation associated with MCT-PAH might imply a putatively higher risk of impaired blood supply in the septal region of the RV cavity. Regretfully, we were unable to experimentally explain the different levels of IKir in SCSMCs.
Implications of lower K+ conductance in SCSMCs
As noted, in addition to the smaller IKir, the voltage-dependent inactivation of Kv appeared to be more sensitive in SCSMCs than that in the other branches in control rats. However, the IBKCa density did not differ between the branches, and no significant changes were observed in MCT-PAH rats (Fig. 5). The SCA used in this study was a branch of the RCA trunk and was dissected from the septal myocardium of the RV. While the RCA and LCA branches were dissected from the surface of the rat heart, the SCA in the relatively deep region might be exposed to higher mechanical stress in the RVH myocardium. Although we currently have no experimental evidence of this activity, it is possible that the putatively different levels of mechanical stress might be associated with the different electrophysiological properties of SCSMCs.
A previous study reported a decrease in right coronary blood flow in PAH patients, and suggested that such a decrease in blood flow could possibly contribute to RVH [10]. Coronary blood flow shows a phasic pattern throughout the cardiac cycle. During systole, the left coronary blood flow is impeded due to squeezing force, but the RCA is relatively much less affected [7]. However, in RVH, the RCA flow becomes biphasic, with reduced systolic and increased diastolic flow. Due to the increase in RV pressure, systolic coronary flow is impeded in PAH patients. The reduced blood flow from the RCA might lead to RV ischemia, which contributes to RV failure. Due to the enlarged RV, the impairment of systolic coronary blood flow in the SCAs might be induced by compression pressure from the LV and RV in PAH with RVH. Accordingly, structural remodeling of the CA wall in RV has been observed in PAH patients and rat models [18]. Although we currently do not have direct evidence, given the anatomical location of the SCAs used in the present study, we suggest that the functional downregulation of IKir in SCSMCs might also occur in the CAs of patients with PAH. The lower Kir and Kv conductance of SCSMCs observed in the present study might be associated with reduced coronary blood flow and CA remodeling in PAH with RVH. Further studies are needed to elucidate the physiological role of K+ currents on blood flow and histological changes in septal CAs.
In summary, this was the first study to comprehensively compare K+ channels in vascular SMCs of CAs from three different branches. The results revealed not only the differential densities of K+ currents, but also branch-specific changes in current amplitudes in PAH-induced RVH. The attenuation of IKir in SCSMCs from a rat MCT-PAH model offered insights into the pathophysiology of the increased risk of RV failure in PAH. Since structural remodeling of the CA wall has been reported in PAH [18], our findings suggest a possible functional remodeling of the CA in PAH-induced RVH.



 XML Download
XML Download