

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Neutrophils are the most abundant circulating human leukocytes essential for protection against pathogens [1]. The bone marrow produces huge numbers of neutrophils, which are released into the circulation during steady-state and inflammatory conditions [2]. During infection, neutrophils are recruited to the site of infection and eliminate pathogens by means of various effector mechanisms, such as the degradation of ingested microbes by fusion of phagosomes with granule proteins, production of reactive oxygen species (ROS) [34567], and release of extracellular chromatin fibers decorated with antimicrobial proteins, termed neutrophil extracellular traps (NETs) [56]. Since these neutrophil effector mechanisms are not only protective but can also be destructive to host tissue, neutrophil production, trafficking, and clearance are tightly regulated by several homeostatic mechanisms in respective organs [7].
Autophagy, an intracellular degradation and energy-recycling mechanism, is one of the key homeostatic process contributing to neutrophil differentiation and function [89]. Mitroulis et al. [10] were the first to discover the evidence of autophagy in neutrophils. Autophagy in neutrophils is activated by phagocytosis of pathogens or activation of pattern recognition receptors (PRRs) by pathogen-derived toxins and molecules [10]. Since the initial discovery of autophagy in neutrophils, many studies have outlined the biological roles of autophagy in neutrophil activities, such as differentiation and effector functions. This review outlines the current understanding of autophagy in neutrophils. We first focus on known autophagy machinery in neutrophils and discuss the role of autophagy in neutrophil differentiation. Finally, we discuss the involvement of autophagy in neutrophil effector functions.
Go to : 
OVERVIEW OF AUTOPHAGY PATHWAYS
Autophagy is an essential mechanism in cell biology. Autophagy is a complex and tightly regulated pathway which allows cells to eliminate damaged or harmful intracellular components to maintain nutrient and energy homeostasis [11]. Hence, autophagy is an important survival mechanism for cells by protecting them from potential harms [12].
Autophagy molecular mechanisms involve the interaction of several conserved autophagy-related (Atg) proteins. Evolutionarily conserved canonical autophagy can be initiated by various stimuli, and results in the formation of a phagophore, whose membrane is derived from the rough endoplasmic reticulum [13141516]. Phagophore formation is initiated by either Atg/ULK1 complex (ULK 1/2-Atg13/FIP200-Atg101) or class III PI3K complex (Atg14-Beclin1/Ambra1-hVPS34/hVPS15), while mammalian target of rapamycin (mTOR) negatively regulates autophagy induction [131416171819]. The phagophore then elongates to form a double-membrane-enclosed autophagosome. During autophagosome formation, two key ubiquitin-like conjugation systems—the Atg12 conjugation system (Atg5-Atg12 in a multimeric complex with Atg16) and the LC3/Atg8 conjugation system (phosphatidylethanolamine [PE]-conjugated LC3-II, which forms after cleavage by Atg4)—are recruited by Atg7 to the autophagosomal membrane [172021]. Conjugated LC3 is integrated into the growing phagophore by the Atg5-Atg12 conjugation system, facilitating growth and elongation of the autophagosome. The Atg5-Atg12 conjugation system dissociates after autophagosome closure and maturation. Alternatively, there is another pathway, a non-canonical autophagy that involves bypass of proteins either in elongation and closure or in initiation in canonical pathway [22]. One example of non-canonical autophagy involves LC3II-associated phagocytosis (LAP). During LAP, a hybrid autophagic-phagocytic process, toll like receptor (TLR)-bound bacteria are taken up into the phagosome. These phagocytosed TLRs triggers the recruitment of LC3II to phagosomal membrane, thereby initiating autophagy and facilitates phagosome maturation [2223]. Then the mature autophagosome fuses with lysosomes. During this final degradative process, the autophagosome undergoes loss of its inner membrane and matures into an autolysosome. This process is initiated by the hVPS34-Beclin complex in association with hVPS38 and requires membrane fusion proteins such as LAMP2/Rab7/SNARES [1820].
Activation of neutrophil autophagic machinery is initiated by various triggers or stimuli, including phagocytosis-dependent and phagocytosis-independent signals [1016]. Such initiating signals induce PI3K activation, leading to enhanced NADPH oxidase (ROS) activity and decreased mTOR signaling [1016242526]. GPCR activation can initiate autophagy in neutrophils via ROS-dependent manner by interaction with high mobility group box 1 (HMGB1) and Beclin-1 [27]. However, macrophage-inducible Ca2+-dependent lectin receptor (Mincle) activation initiates autophagy in neutrophils independent of ROS and mTOR manner through activation of Beclin-1 nucleation complex [28]. Neutrophil nutrient starvation can also initiate autophagy via the AMPK pathway [2329]. Nucleation processes mediated by ULK (inhibition of mTOR) and/or Beclin-1 are followed by recruitment of other autophagy proteins to the phagosome, resulting in autophagosome formation [172021]. Ubiquitin-like conjugation system components Atg5 and Atg7 are key regulators of phagophore elongation and autophagosome formation [172021]. Atg7 induces LC3 lipidation, whereas the Atg12-Atg5-Atg16 complex integrates LC3-II into the phagophore and completes autophagosome formation. Knockdown of these genes inhibits autophagy in neutrophils [3031]. We have also previously reported that defective Atg5-Atg12 complex formation compromises phagophore elongation, resulting in defective autophagy [32]. In addition, p62/SQSTM is incorporated into the completed autophagosome and undergoes degradation in the autolysosome [33]. Increased p62/SQSTM degradation is strong indirect evidence that autophagy is occurring [33], and decreased p62/SQSTM1 degradation is associated with defective autophagy or inhibition of autophagy machinery [323435].
In summary, autophagy is initiated by various stimuli which either induce activation of Beclin-1 or inhibition of the mTOR pathway. These events then induce phagophore nucleation and elongation by the Atg5-Atg12 complex. Finally, autophagosome formation is completed by incorporation of lipidated LC3-II into the phagosome membrane, and p62/SQTM1 is degraded (Fig. 1). Similarly, activation or regulation of neutrophil autophagy follows autophagy machinery as discussed above. Neutrophil can sequester materials by mTOR-ULK1 dependent canonical autophagy or ATG5/ATG7 dependent non-canonical pathway with LC3 associated structures (LAP). While all stimuli which initiate autophagy in neutrophils up-regulate LC3 expression and lipidation, differential expression of ATG genes (ATG3, ATG4, ATG6 and ATG7) is observed depending on the stimuli [10]. Therefore, monitoring expression levels of Beclin-1 (Atg6), Atg5, Atg12, and p62/SQSTM1, along with LC3-II (LC3 expression and lipidation) can provide improved insights into the differential induction and regulation of autophagy machinery in neutrophils.
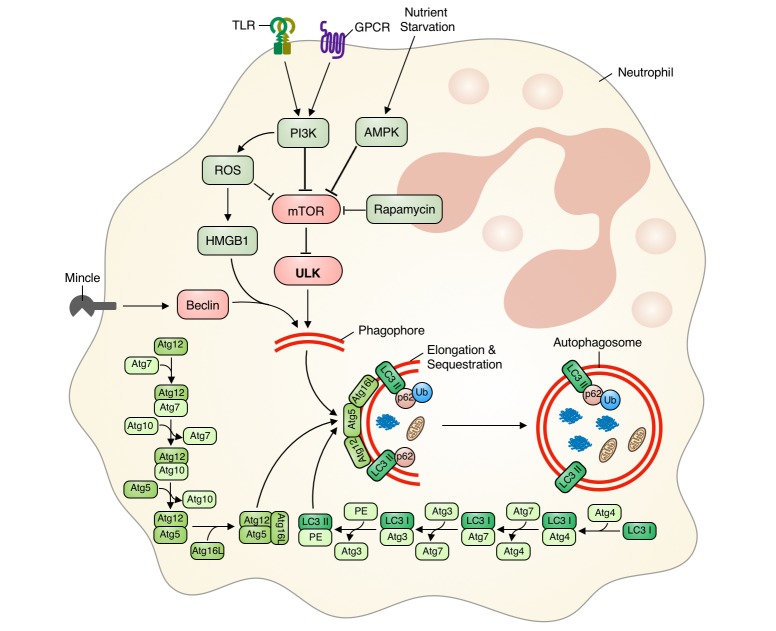 | Fig. 1Regulation of autophagy in neutrophils.Phagocytosis-independent autophagy in neutrophils is initiated by starvation or activation of receptor by different stimuli. This induces either Beclin-1 activation or mTOR inhibition via increased ROS, PI3K, and/or AMPK activity. Activation of Beclin-1 or mTOR inhibition induces phagophore nucleation and elongation by the Atg5-Atg12 complex. Finally, autophagosome maturation is completed via incorporation of lipidated LC3-II into the phagosome membrane, and the ubiquitin protein p62/SQTM1 is degraded. mTOR, mammalian target of rapamycin; ROS, reactive oxygen species; TLR, toll like receptor.
|
Go to :
AUTOPHAGY IN NEUTROPHIL DIFFERENTIATION
Neutrophils are generated from hematopoietic precursors in the bone marrow. Hematopoietic stem cells differentiate into common myeloid progenitor cells, which further differentiate into neutrophil precursors. These successive stages of neutrophil differentiation are governed by transcriptional programs [3637]. Although many reports discuss the role of autophagy in hematopoiesis and hematopoietic cell fate, its role in neutrophil differentiation has not been fully understood. Selective autophagy, macrophagy and mitophagy, differentially regulates the differentiation and maturation of myeloid lineage cells such as granulocytes, erythrocytes etc. [383940]. However, impaired autophagy in myeloid cells leads to hematopoietic diseases such as acute lymphoid leukemia [39]. Once committed to the granulocyte lineage, these progenitor cells pass through a mitotic pool composed of myeloblasts and promyelocytes. They further differentiate into myelocytes, metamyelocytes, band cells, and, finally, into mature segmented neutrophils [41]. There is increased autophagic activity in myleoblasts, promyleocytes and myleocytes, and declined as it differentiates into metamyleocytes and band cells. Autophagy is lowest in cells that had undergone terminal differentiation [3442]. Lipophagy, the autophagic ability to degrade fatty acid-enriched lipid droplets provides free fatty acids, the substrate, for oxidative phosphorylation to enhance ATP generation required for differentiation. Knockdown of either ATG5 or ATG7 results in defective neutrophil precursors differentiation into promyelocytes [42]. Such autophagy-defective neutrophil precursors exhibited an impaired lipophagy, mitochondrial respiration and ATP production, with a concomitant increase in glycolytic activity. As fatty acids represent an essential substrate for oxidative phosphorylation during the early stages of neutrophil differentiation, the resultant lack of ATP impairs differentiation during these stages [42]. However, tissue specific downregulation of ATG5 expression in immature neutrophils resulted in elevated proliferation of neutrophil precursor in bone marrow and an accelerated process of neutrophil differentiation. These ATG5 deficient neutrophils were without any abnormalities in morphology, granule protein content, apoptosis regulation and other effector functions. But, as a consequence of reduced autophagy (possibly mitophagy) the number of mitochondria were increased but did not affect the neutrophil morphology and function [34]. Moreover, over-expression of autophagy by overexpression of ATG5 or using pharmalogical approaches, delayed terminal neutrophil differentiation [34]. Overall, autophagy is upregulated in myeloblasts and progressively down-regulates as they continue to differentiate. These results highlight the important role of autophagy during early mitotic stage of neutrophil differentiation and possibly due to short life span, active autophagy is not important for post-mitotic stage of neutrophil differentiation. Altogether, autophagy appears to reciprocally regulate neutrophil differentiation in mitotic and post-mitotic pools.
Go to :
AUTOPHAGY IN NEUTROPHIL BACTERICIDAL ACTIVITY
Neutrophils are professional phagocytes and are thus able to phagocytose and destroy infectious agents [43]. During phagocytosis, pathogens become sequestered in highly-organized endocytic compartments known as phagosomes, and are ultimately degraded via fusion of these compartments to lysosomes [44]. During phagocytosis by neutrophil, bacteria is engulfed into phagosomes. Fusion of granule contents and NADPH complex to phagosome contribute to its maturation and antimicrobial activity [45]. Recent studies indicate neutrophils exhibits phagocytosis-dependent or non-canonical selective autophagy during bactericidal activity [264647]. During phagocytosis dependent LAP, PRR-bound bacteria in phagosome induces LC3 translocation, thereby initiating autophagy. LAP facilitates phagosome maturation and consequent bacteria degradation [23]. Phagocytosisindependent autophagy (or non-canonical selective autophagy, xenophagy) also exists, and is initiated by ubiquitinated p62/SQSTM1 bound to intracellular bacteria or bacteria which have escaped the phagocytic pathway. Such autophagy-mediated bacterial clearance is rapid and occurs independently of ROS generation [48]. Furthermore, augmentation of autophagy greatly enhanced neutrophil phagocytosis of bacteria and its bactericidal activity, facilitating effective elimination of both drug-sensitive and multi drug-resistant bacterial strains [234950]. In contrast, impaired or inhibited autophagy enhanced survival and growth of bacteria within neutrophils [235152]. These findings indicate the important contribution of non-canonical selective autophagy to the bactericidal activity of human neutrophils and possibly the defect with these processes rather than canonical pathway may hinder bacterial clearance.
Go to :
AUTOPHAGY AND NETosis
Entrapment and killing of pathogens is also mediated by NETosis (a type of cell death culminating in NETs release), first discovered by Brinkmann et al. [5] as a novel extracellular microbicidal mechanism employed by neutrophils. NETs are composed of dense extracellular chromatin studded with antimicrobial proteins (e.g., histones, neutrophil-derived granule proteins, and other cytosolic proteins) [5354]. During NETosis, chromatin becomes decondensed, followed by nuclear swelling, nuclear membrane disintegration, rupture of the cell membrane, and release of NETs into the extracellular space. Released NETs are eventually phagocytosed and eliminated from circulation by macrophages [55].
Various stimuli induce neutrophil NETs formation, including pathogen associated molecular patterns (e.g., Staphylococcus aureus, Candida albicans, Leishmania spp., and other bacteria and fungi), exogenous compounds (e.g., phorbol myristate acetate, calcium ionophores, and potassium ionophores), danger associated molecular patterns (e.g., interleukin [IL]-33, immune complexes, auto-antibodies, histones, LL-37, and HMGB1), inflammatory cytokines (e.g., tumor necrosis factor-α and IL-8), platelets, and antibodies [565758596061]. Different stimuli induce activation of differential signaling pathways which nonetheless converge on NETosis [566062].
Remijsen et al. [63] reported that both autophagy and ROS are required for NETosis. They demonstrated that both autophagy and ROS generation are required for chromatin decondensation: defects in either pathway result in apoptotic cell death rather than NETosis [63]. Although this suggests that the ROS-autophagy axis is essential for NETosis, autophagy can be induced in both ROS-dependent and ROS-independent manners [63]. Calcium ionophore induces rapid, NADPH oxidase-independent NETosis [64], and NETosis induced by certain bacteria is known to be NADPH oxidase-independent [656667]. Interestingly, Itakura and McCarty described regulation of autophagy and cytoskeletal remodeling by the mTOR pathway during NETs formation [16]. Inhibition of mTOR or autophagy activation by rapamycin enhanced NETs formation, whereas autophagy inhibition by wortmannin (a PI3K inhibitor) diminished NETs release [16]. Since autophagy is reciprocally regulated by the mTOR pathway, this study indicates that ROS-independent NETosis is mediated by mTOR-autophagy signaling. Moreover, evidence is accumulating that autophagy is an important regulator of NETs formation. Chargui et al. [23] observed that autophagy preceded NETs formation in neutrophils infected with invasive Escherichia coli, and that Atg5 silencing completely blocked NETs formation. Pharmacological inhibition of autophagy (using wortmannin, 3-MA, bafilomycin, chloroquine, or atrazine) or inhibition of autophagy via siRNA-mediated ATG7 silencing both inhibited NETs release [2832616869]. Neutrophils isolated from aged mice possessing a defect in ATG5 (and thus in autophagic pathways) exhibited decreased NETs release [30]. Additionally, Mincle knockdown attenuated autophagy, concomitant with impaired NETs formation [28]. We have previously observed that impaired autophagy in neutrophils results in decreased NETosis during sepsis [32]. Furthermore, pharmacological induction of autophagy in neutrophils (using rapamycin or tamoxifen) rescued NETs formation [2832]. However, Germic et al. [70] reported that neutrophils isolated from ATG5 -knockout mice still retained the capacity for NETs release. Furthermore, only PI3K inhibition attenuated NETs formation, while inhibition of downstream autophagy pathways (using bafilomycin or chloroquine) did not.
Further, autophagy and NETosis are closely related with various pathologies, both in experimental disease models and in the clinical conditions [31717273]. Autophagy positively regulates NETosis, and impaired autophagy thus correlates with reduced NETs formation [283263]. Decreased NETosis and resultant prolonged neutrophil survival increases mortality among sepsis patients [74]. Furthermore, NETs depletion resulted in a delayed immune response and polymicrobial sepsis in mice [75]. We previously demonstrated that neutrophils isolated from patients experiencing fatal sepsis exhibit decreased NETs formation. Additionally, decreased NETs formation correlated with impaired autophagy, and autophagy augmentation protected mice from sepsis in a NETs-dependent manner [33]. Furthermore, clarithromycin, a macrolide antibiotic, induced autophagy-dependent formation of NETs decorated with the antimicrobial peptide LL-37. Clarithromycin-induced NETs formation inhibited growth and biofilm formation of Acinetobacter baumannii [76]. Tamoxifen, an autophagy inducer, restored impaired autophagy and NETs formation in Mincle-defective neutrophils and enhanced neutrophil phagocytosis of Klebsiella pneumoniae, limiting bacterial pneumonia [77]. One primary function of NETs is sequestration of inflammatory sites from circulation to prevent pathogen spreading. However, tumor-infiltrating neutrophils also exhibited increased autophagy and NETs formation. Such NETs may physically sequester cancer cells, interfering with the anti-tumoral activities of effector T cells [7879]. Moreover, cancer cells exhibit increased metastatic rates in a NETs-enriched environment [808182]. These studies showed autophagy is important but not essential for NETosis. However, as discussed above intervention of both canonical and non-canonical autophagy with molecular or pharmacological interventions impacts NETsosis. Dissecting among the ATG-dependent autophagy, ATG-independent autophagy and ATG-dependent non-autophagic pathway during NETosis may further clarify the importance of autophagy during NETosis.
Go to :
AUTOPHAGY AND NEUTROPHIL GRANULE FORMATION/DEGRANULATION
Neutrophils possess various cytoplasmic granules: azurophil (primary) granules, specific (secondary) granules, gelatinase-containing tertiary granules, and secretory vesicles (phosphasomes) [838485]. A recent study also described a fourth (ficolin-1-rich) neutrophil granule [71]. Granule heterogeneity is explained by a targeting-by-time hypothesis: granules are formed and stored in a time-dependent order which corresponds to neutrophil developmental stages. Thus, granule formation occurs throughout neutrophil development, beginning with azurophil granules (in myeloblasts-promyelocytes) and ending with secretory vesicles (in band cells) [718384]. Different transcription factors regulate expression of different granule proteins [71]. Recent studies have demonstrated the involvement of autophagy in granule formation. Knockdown of ATG7 correlated with decreased expression of the transcription factor C/EBPε (involved in myeloid differentiation and granule maturation), leading to defective neutrophil granule maturation and fewer neutrophil granules with diminished content [42]. Such neutrophils exhibited increased primary and secondary granule protein expression, but decreased tertiary granule protein expression. Since tertiary granules are formed during the band cell stage, while primary and secondary granules are continually produced and degraded throughout neutrophil differentiation [72738486], it is speculated that autophagy might be involved in regulation of the relevant transcription program governing granule protein formation prior to the band cell stage.
Autophagy is also involved in neutrophil degranulation. Activated neutrophils release their granule contents into the phagosome and/or extracellular space via exocytosis, in a process known as degranulation [87]. Degranulation occurs in an order opposite to that of granule formation, with secretory vesicle contents being released first and azurophil granule contents being released last [738384]. Knockdown of ATG5 and ATG7 led to impaired ROS generation and impaired degranulation, reducing inflammatory function of neutrophils [31]. Autophagy deficiency diminished neutrophil degranulation (primary and secondary granules), with the exception of myeloperoxidase (MPO) [31]. Since the total intracellular quantity of neutrophil MPO remained consistent regardless of Atg knockdown [4283], the above phenomenon is likely due to storage impairment of MPO in Atg-knockdown neutrophils. These studies suggest a role for both conventional secretion and selective autophagy in neutrophil granule production, storage, and content release. Further dissecting the molecular signatures of autophagic process may facilitates understanding the specific process associated with neutrophil granule formation and degranulation.
Autophagy is also involved in the production and release of certain neutrophil cytokines. Neutrophils produce pro- and anti-inflammatory cytokines either spontaneously or in response to stimulation [88899091]. The pro-inflammatory cytokine IL-1β is one of the most extensively studied neutrophil-derived cytokine [929394]. Autophagy is involved in the release of IL-1β by human neutrophils in an ATG-dependent secretory pathway. Neutrophil release of IL-1β was inhibited by autophagy inhibitors or ATG5 silencing [94]. This is particularly interesting given that autophagy degrades and blocks maturation and secretion of IL-1β in other immune cells such as macrophages [9596].
Go to :
AUTOPHAGY AND NEUTROPHIL-MEDIATED INFLAMMATION/PATHOLOGY
Autophagy is involved in modulation of inflammation through regulating pathogen clearance, antigen presentation, cytokine generation, and immune responses [97]. Abnormalities in autophagic machinery have been linked to several diseases, such as Crohn's disease, cystic fibrosis, and inflammation-associated metabolic disease [98]. As we have reviewed, autophagy has also been established as a regulator of neutrophil function [31]. Excessive or dysregulated neutrophil functions has been noted in various diseases such as sepsis, chronic obstructive pulmonary disease (COPD), chronic granulomatous disease, Crohn's disease, and familial Mediterranean fever (FMF) [99100101]. Recently, the role of autophagy in neutrophil-mediated inflammation and autoimmune disease has been established [31]. Inhibition of autophagy during neutrophil-mediated inflammation and autoimmune disease reduced disease severity and progression by suppressing degranulation and ROS production [2335]. Similarly, suppression of autophagy through NLRP3 knockdown or inhibition of the NLRP3 inflammasome enhanced neutrophil recruitment and phagocytosis, thereby improving bacterial clearance and augmenting the survival of septic mice [102]. Furthermore, during the pathogenesis of COPD, autophagy induction increased neutrophil apoptosis and accelerated disease development via platelet-activating factor receptor (PAFR) [27]. Inhibition of PAFR-dependent autophagy reversed autophagic neutrophil death, and slowed disease development [27]. In contrast, autophagy defects in myeloid cells attenuated neutrophilic differentiation of acute promyelocytic leukemia cells and lowered the efficacy of all-trans retinoic acid therapy [103104105]. Inhibiting autophagy in myeloid cells also resulted in spontaneous lung inflammation and enhanced susceptibility to endotoxemia, an effect attributed to increased levels of IL-8 [106]. Similarly, adherent-invasive E. coli (AIEC)-infected neutrophils exhibited a defect in the degradation stage of autophagy (delayed or limited autolysosome maturation), enhancing inflammation via autophagic neutrophil death and increased IL-18 secretion [23]. Further, neutrophil ATG5 knockdown induced dysregulated function, increasing the susceptibility to M. tuberculosis infection [35]. In contrast, autophagy activation through serum starvation or rapamycin treatment of neutrophils enhanced their bactericidal activity against AIEC [23]. Autophagy augmentation in neutrophils by intravenous immunoglobulin treatment also enhanced bactericidal activity against multi-drug resistant strains [49]. However, increased autophagy in neutrophils is associated with IL-β-mediated inflammatory responses during active ulcerative colitis and FMF [9293107]. This is attributed to the increased load of IL-1β in autophagydriven NETs. Increased IL-1β-induced REDD1 (regulated in development and DNA damage response 1) expression enhances autophagy in neutrophils, leading to increased formation of NETs loaded with IL-1β [9293107].
These studies suggest a dual role for autophagy in neutrophil function during inflammation (Fig. 2). Augmentation of autophagy may be an effective target for enhancement of proper myeloid differentiation and antimicrobial defense. Conversely, autophagy inhibition may be useful in neutrophil-mediated inflammatory disease. Broadly, autophagy is important for neutrophil differentiation and function. However, one important question is whether these pathological outcomes are mediated by canonical or non-canonical autophagic processes. This implies that understanding the molecular mechanism of autophagy during cell physiology and pathological condition would further clarify role of specific autophagy during infection and inflammation.
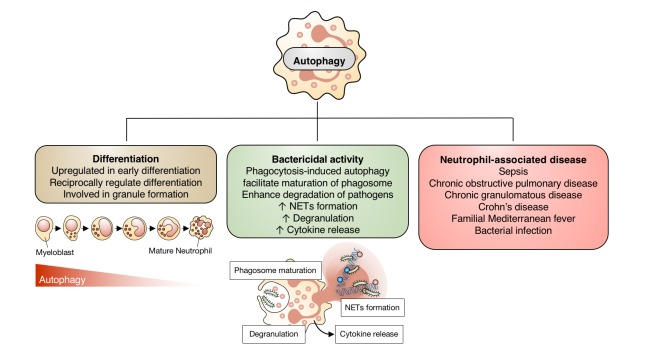 | Fig. 2Contribution of autophagy to neutrophil function.Autophagy is involved in neutrophil differentiation and granule formation. It enhances neutrophil phagocytosis, neutrophil extracellular traps (NETs) formation, degranulation, and cytokine release. Autophagy in neutrophils also differentially modulates the outcomes of inflammation and infection in a context-dependent manner.
|
Go to :
CONCLUSIONS
Autophagy is an important phenomenon involved in sequestering cytoplasmic materials and pathogens. Autophagy in neutrophil is associated with its differentiation (lipophagy) during early stage. Autophagy is associated with regulation of degranulation and cytokine production (secretory autophagy), bactericidal activity (xenophagy and LAP) and NETs release. Depending on the inflammatory environment, autophagy in neutrophil may behave as a double-edged sword. While it is beneficial in combating infection, it may favor excessive inflammation through exaggerated NETs formation and cytokine release. Thus, Autophagic homeostasis is important for proper neutrophil effector function and host health. Learning to distinguish between canonical and non-canonical autophagic process could add a significant contribution to understanding autophagy regulation of neutrophil's fate and function. This essential insight may also contribute towards appropriate development of novel pharmacological therapies targeting autophagy in neutrophil-mediated diseases.
Go to :
 XML Download
XML Download