

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Drug reaction with eosinophilia and systemic symptoms (DRESS) syndrome is a serious cutaneous adverse event associated with constitutional symptoms [1]. Flare-up reactions can occur as a continuum from initial drug-related immune reactions. However, they can also present as a transformed T-cell-mediated hypersensitivity reaction after a patient recover from the initial manifestations. The mechanisms underlying flare-up reactions are difficult to understand because they are multifactorial and involve complex immunological components, particularly reactivation of latent viruses [23]. Currently, flare-up of DRESS is considered to be an immune reconstitution inflammatory syndrome (IRIS) associated with either discontinuation or a reduction in the dose of immunosuppressive agents [3].
CASE REPORT
A 60-year-old man with chronic kidney disease was referred to an allergy clinic for a second opinion regarding waxing/waning of the condition accompanied by generalized erythroderma for 6 months. Ten months prior to referral, he was prescribed allopurinol to treat hyperuricemia. After 3 months of treatment, he suffered a generalized maculopapular skin eruption, high fever, eosinophilia, and elevated liver enzymes. Analysis of human leukocyte antigens revealed that he harbored the HLA-B*58:01 allele, a risk marker for allopurinol-induced DRESS [4]. Allopurinol was withdrawn immediately and replaced with a systemic corticosteroid equivalent to prednisolone (1 mg/kg), along with other supportive measures. After administration of systemic corticosteroid, the rash improved gradually, and liver function and eosinophil counts returned to normal. However, he was admitted to a hospital several times due to repeated episodes of generalized eruption, which eventually manifested as erythroderma accompanied by fever, chill, facial swelling, pedal edema, and inguinal lymphadenopathy. Several attempts at steroid reduction resulted in generalized eruption followed by widespread skin exfoliation; these symptoms improved when the steroid dose was increased. Laboratory findings revealed a white blood cell count of 6.67×103/mm3 (eosinophils, 14%), a C-reactive protein level of 1.97 mg/dL (normal range, 0–0.5 mg/dL), serum creatinine value of 2.07 mg/dL (normal range, 0.5–1.2 mg/dL), and aspartate transaminase/alanine transaminase level of 15/7 IU/L. Other laboratory indexes and electrocardiogram were within normal range. Peripheral blood analysis revealed a very high load of cytomegalovirus (CMV; 10,375 copies/mL) and Epstein-Barr virus (EBV; 132,988 copies/mL). Human immunodeficiency virus (HIV) and human herpesvirus 6 (HHV-6) were not detected, but he was positive for HHV-8 (Table 1). A computed tomography scan revealed enlargement of multiple lymph nodes in the axillary, mediastinal, para-aortic, retrocaval, and aortocaval areas. A skin biopsy from the dorsum of a foot showing erythroderma revealed acanthosis with superficial perivascular lymphohistiocytic and mild eosinophilic infiltration; the tissue stained positive for HHV-8. Analysis of inguinal lymph nodes revealed an atypical vascular proliferative lesion that was positive for EBV and HHV-8; these findings are consistent with Kaposi sarcoma and ruled out the possibility of lymphoma.
Table 1
Laboratory test of the patient with DRESS syndrome

DRESS, drug reaction with eosinophilia and systemic symptoms; WBC, white blood cell; CRP, C-reactive protein; CMV, cytomegalovirus; EBV, Epstein-Barr virus; HIV, human immunodeficiency virus; HHV, human herpesvirus.
![]()
Peripheral blood mononuclear cells (PBMCs) were isolated from the patient and from three age- and sex-matched controls. Flow cytometry analysis showed that the percentage of CD3+ T cells in the patient was higher (Fig. 1A, B) than that in controls, with a marked increase in the percentages of T cells positive for GATA3 or FOXP3, which are transcription factors representative of Th2 and regulatory T (Treg) cells, respectively. By contrast, the percentage of T-bet-positive cells, which is representative of Th1 cells, was lower in the patient than in controls (Fig. 1C, D). In parallel with this, T cells isolated from the patient showed increased production of type 2 cytokines such as interleukin (IL)-5 and IL-13, but reduced production of interferon (IFN)-γ, upon PMA/ionomycin stimulation in vitro (Fig 1E, F). Simultaneously, IL-13+ T cells from the patient showed increased production of IL-17. In vitro stimulation studies confirmed the role of the viruses detected previously. When PBMCs were stimulated with peptides derived from CMV or EBV, they secreted more IL-4 and IL-13 into the culture supernatant; however, the amount of IFN-γ remained unchanged (Fig. 1G). Next, we examined the distribution of DCs and monocytes that activate T cells by presenting antigen. Dendritic cells (CD11c+HLA-DR+) and classical monocytes (CD11b+CD14+CD16-) within the PBMC population isolated from the patient with DRESS were markedly depleted (Fig. 2A, B). By contrast, the population of nonclassical monocytes (CD11b+CD14-CD16+) expressing costimulatory molecules (CD40, CD86) was elevated (Fig. 2A-C).
 | Fig. 1Flow cytometry analysis of lymphocyte subpopulations within the peripheral blood mononuclear cell (PBMC) population. (A) Flow cytometry analysis of B cell (CD45+CD3-CD19+) and T cell (CD45+CD3+CD19-) subsets within the PBMC population of normal controls (n=3) and the patient. Values in the diagrams indicate the percentage of the gated population. (B) Comparison of B cell, T cell, CD4+ T cell, and CD8+ T-cell populations within PBMCs from normal controls and the patient. (C) Flow cytometry analysis of transcription factor expression by T cells. (D) Expression of T-bet, GATA3, RORγt, and Foxp3 by T cells. (E) Cytokines produced by T cells, as analyzed by intracellular staining and flow cytometry. (F) Comparison of cytokine production by the cells in (E). (G) Comparison of cytokine levels in medium from PBMCs (from the controls and the patient) cultured with CMV or EBV peptides or allopurinol. Data are expressed as the mean±standard deviation. DRESS, drug reaction with eosinophilia and systemic symptoms; IFN, interferon; IL, interleukin; CMV, cytomegalovirus; EBV, Epstein-Barr virus.
|
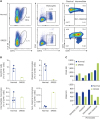 | Fig. 2Comparison of mononuclear phagocyte populations within PBMCs. (A) Distribution of dendritic cells (CD45+CD15-CD11c+HLA-DR+) and monocytes (CD45+CD15-CD11b+) and their subsets within the PBMC population from normal controls (n=3) and the patient. Values in the diagrams indicate the percentage of the gated population. (B) Comparison of dendritic cell and monocyte subsets, including classical (CD45+CD15-CD11b+CD14+CD16-) monocytes, intermediate (CD45+CD15-CD11b+CD14+CD16+) monocytes, and nonclassical (CD45+CD15-CD11b+CD14-CD16+) monocytes, within PBMCs. (C) Mean fluorescence intensity of CD40 and CD86 expressed by dendritic cells and each subset of monocytes. Data are expressed as the mean±standard deviation. PBMC, peripheral blood mononuclear cell; DRESS, drug reaction with eosinophilia and systemic symptoms.
|
DISCUSSION
IRIS following withdrawal of immunosuppressive medications has been reported in both non-HIV patients and HIV-infected patients [5]. The clinical correlation between high viral load and severity of constitutional symptoms suggests that viral reactivation could be responsible for aggravating IRIS symptoms. It is proposed that drug metabolite-induced hypogammaglobulinemia and B-cell depletion in DRESS reactivate latent viruses such as HHV-6, EBV, and CMV [6]. Our result demonstrated that IL-13+ T cells may be generated in response to reactivation of EBV and CMV following a reduction in B-cell counts and immunoglobulin production (data not shown).
DRESS syndrome is induced by drug-specific T cells and corresponds to a type IV hypersensitivity with eosinophilic activation; in addition, Th2-type immune responses may be involved [7]. Here, we found that eosinophilia occurred in the skin of the patient with DRESS syndrome, and that the percentages of IL-5- and IL-13-producing T cells within the PBMC population were increased, as reported previously [8]. Moreover, there was an increase in the percentage of IL-17-expressing IL-13+ T cells within the PBMC population from the patient with DRESS, a phenotype observed in cases with severe allergic responses [9]. However, it is not clear how a small compound such as a drug activates T cells and promotes cytokine production. There are several hypotheses about the mechanism underlying T-cell activation in DRESS. Pichler [10] suggested that drugs or their metabolites act as a haptens that bind to autologous proteins and induce cellular or humoral immune responses. Shiohara et al. [11] suggested that pre-existing viral-specific T cells might cross-react with specific drugs. In addition, Criado et al. [12] reported that the viral reactivation observed in IRIS might provide danger signals to nonspecific T cells, which then induce oligoclonal expansion of drug-specific and viral-specific T cells. Treg cells are likely to play an important role in IRIS in non-HIV patients, but their role in DRESS is not completely understood [35]. During the acute phase of DRESS, Treg cells expand and possibly inhibit viral-specific immune responses, leading to viral reactivation. During the resolution phase, Treg cells lose their normal function, which may be linked to hypersensitivity to drugs and flare-up reactions [13]. In line with previous reports, our data show an increased Treg population in the DRESS patient who developed a prolonged course of IRIS. However, it is unclear whether these Treg cells were functionally intact and responsible for suppressing antiviral immune responses in this patient. Therefore, additional experiments are needed to validate the function of Treg cells.
We found a clear distinction between mononuclear cells (monocytes and dendritic cells) that play an important role in tissue homeostasis and those that elicit an immune response to invading pathogens. Under normal conditions, viral infection induces rapid differentiation of monocytes into dendritic cells [14]. However, dendritic cells were depleted from the patient with DRESS. It is noteworthy that the number of nonclassical monocytes showing high expression of costimulatory molecules was increased, while that of classical monocytes was depleted severely. Monocytes are a highly plastic and heterogeneous population of cells showing different functional phenotypes in response to environmental change. Most human monocytes (~85%–90%) are classical monocytes; the remaining ~10%–15% are further subdivided into intermediate and nonclassical subsets [15]. Classical monocytes traffic to sites of inflammation and play a major role in phagocytosis, as demonstrated by high peroxidase activity, high levels of IL-10, and low levels of tumor necrosis factor (TNF)-α in response to lipopolysaccharide (LPS) stimulation [16]. Intermediate monocytes are a transitional population that bridges the classical and nonclassical monocyte subsets and exhibits pro-inflammatory functions, including production of TNF-α and IL-1β upon stimulation by LPS [16]. Intermediate monocytes not only display both phagocytic and inflammatory functions, but also play a role in the treatment of patients with rheumatoid arthritis [1718]. Nonclassical monocytes show the strongest pro-inflammatory responses to TLR stimulation [19]. Also, CD14-CD16high monocytes in peripheral blood of DRESS patients act as precursors that show increased expression of CCR4, a skin-homing chemokine receptor; indeed, these cells migrate to the skin, present viral particles to CD4+ T cells, and induce drug allergy responses [20]. Previous reports of patients with sepsis and systemic lupus erythematosus demonstrated that nonclassical monocytes also displayed properties characteristic of antigen presenting cells, and expressed inflammatory phenotype under higher levels of pro-inflammatory cytokines and lower levels of anti-inflammatory IL-10 [18]. In the current case, virus-induced TLR7/9 signaling may contribute to activation of human nonclassical monocytes and to persistence of inflammatory responses through high expression of costimulating molecules such as CD40 and CD86, which activate T cells. However, further studies are warranted to elucidate whether these nonclassical monocytes play a role in the persistent virus-associated flare-up reactions in IRIS.
Taken together, the data presented herein suggest that skewed subsets of T cells, B cells, and monocytes are responsible for strengthened Th2 inflammatory responses and vulnerability to viral reactivation in a DRESS patient who exhibited prolonged flare-ups and persistent viral reactivation despite long-term withdrawal of the culprit drug. Therefore, patients with prolonged flare-up of DRESS should be evaluated carefully with respect to associated viral reactivation and related alterations in lymphocyte and monocyte populations. Moreover, we look forward to new advances in understanding the interplay between lymphocytes and monocyte populations, and to identifying potential therapeutic targets that facilitate control of prolonged flare-up reactions in DRESS.
 XML Download
XML Download