

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Buffalo (Bubalus bubalis) is an important livestock in the animal husbandry ecosystem in South China and presents a high feed conversion rate, high nutritional value of its dairy products and resistance to crude feed. Buffalo also have some disadvantages, such as their low breeding efficiency, long cycle required to improve their varieties and fewer high-quality buffaloes in China compared with other countries. To solve these problems, researchers have used various reproductive biotechnologies [1], including artificial insemination [2], intracytoplasmic sperm injection [3] and in vitro fertilization [4]. In recent years, research on spermatogonial stem cells (SSCs) has attracted substantial attention. SSCs located on the basement membrane of the seminiferous tubules [5], are the precursor cells of sperm, providing a continual spermatogenesis process and thereby ensuring the transfer of genetic material from parent to offspring.
Thus far, considerable research progress has been made on the in vitro culture of SSCs. The commonly used methods for in vitro SSC culture include culture systems with undefined (such as foetal bovine serum [FBS]) [67891011121314] and defined (such as KnockOut Serum Replacement [KSR] and bovine serum albumin [BSA]) materials [1516171819202122]. Studies have shown that using KSR instead of FBS could effectively inhibit the differentiation of male germ cells in mouse testis [16]. When culturing immature SSC-like cells of rat testis tissue, the culture effect of KSR was better than that of FBS [17]. Compared with FBS, KSR could significantly increase the efficiency of clone formation and self-renewal of bovine SSCs [19].
Most studies on buffalo SSCs have used culture systems with undefined materials, while some have also attempted to culture buffalo SSCs in systems using defined materials in recent years [232425]. However, the effects of different culture systems on buffalo SSCs were still inconclusive. In this study, we compared the effects of culture systems with undefined materials and culture system with defined materials on the in vitro culture of buffalo SSC-like cells. As a special type of adult stem cell, SSCs have the molecular characteristics of both stem cells and germ cells; thus, we used the undifferentiated SSC-like cell marker UCHL1 [26], the marker of inchoate buffalo SSC-like cells NANOS2 (nanos C2HC-type zinc finger 2) [27] and the germ cell marker DDX4 to comprehensively evaluate the cells we obtained [26].
MATERIALS AND METHODS
Reagents and animal ethics
All reagents used in this study were purchased from Sigma-Aldrich Company (USA) unless otherwise stated. All animal procedures used in this study were approved by the Animal Care & Welfare Committee of Guangxi University.
Collection of buffalo testis
The buffalo testes (3 pairs, 3- to 6-months) were collected from the Animal Experiment Center of Guangxi University (animal study approval number: GXU2016-017). The testes were kept in Iscove's Modified Dulbecco's Medium (IMDM, Gibco, USA) containing penicillin-streptomycin (100 U; ThermoFisher Scientific, USA) and transported on ice to the laboratory within 2 h.
Separation and enrichment of cells from buffalo testis
Cells were isolated from the buffalo testis as follows: the testis was sterilized and washed with 75% alcohol and phosphate buffered saline (PBS), and the tunica albuginea was then removed. Next, the tissue was cut into the smallest possible pieces and incubated in IMDM containing collagenase IV and DNase I (Worthington Biochemical Corp., China) at 37°C for 40 min to promote digestion into fragments. These fragments were centrifuged in IMDM at 2,000 rpm for 5 min, resuspended with PBS, and centrifuged twice at 2,000 rpm for 5 min. Next, the seminiferous fragments were incubated in IMDM containing 0.25% trypsinase (Gibco) and DNase I for 3 to 5 min for single-cell digestion. Then, the digestion was halted with IMDM containing 10% FBS (HyClone, USA). The debris was removed using nylon cell strainers (BD Falcon, USA).
The differential plating method was used to enrich the SSC-like cells. The single-cell suspension was seeded in a dish coated with 0.2% gelatine and cultured in IMDM containing 10% FBS at 37°C for 24 h. Thereafter, the adherent cells, referred to as G-cells, that were adsorbed onto the gelatine were collected, seeded in a dish containing 0.01% Collagen I protein and cultured in IMDM containing 10% FBS at 37°C for 4 h to remove the remaining somatic cells and differentiated germ cells. Then, the nonadherent cells, referred to as C-cells, that were adsorbed onto the Collagen I protein were collected, seeded in a dish coated with laminin (18 μg/μL), which could specifically adsorb the SSC-like cells, and cultured for 40 min. The nonadherent cells were discarded, and 0.5% (w/v) BSA-PBS was then used to elute the bound cells. The final obtained cells were referred to as L-cells and were used in the subsequent experiments.
Immunocytochemistry staining of SSC-like cells by the cell smear method
L-cells were fixed with 4% paraformaldehyde at room temperature and centrifuged at 2,000 rpm. The supernatant was then discarded, and the cell pellet was resuspended in 1 mL of PBS. A drop of the cell suspension was added to an adhesive slide, and the side of a pipette tip was used to smear the cells. The slides were placed in a dry incubator to evaporate the liquid. Then, the cells were blocked using a blocking solution containing 2.5% horse serum for 1 h at room temperature. The slides were then incubated with primary antibodies against DDX4 (1:200; Abcam, USA) and UCHL1 (1:200; Abd Serotec, UK) for 1 h at room temperature and washed with PBS++ (BOSTER, China) 3 times for 5 min each. Subsequently, the slides were incubated with a goat anti-rabbit IgG H&L (FITC) secondary antibody (1:500; Abcam) for 1 h at room temperature and washed with PBS++ 3 times for 5 min each time. A drop of DAPI was placed onto each slide, which was then sealed with a coverslip. Fluorescence images were captured with the NIS Elements imaging system (Olympus TH4-200; Olympus, Japan) and processed and analysed with Photoshop CS5 software (Adobe Systems Inc., USA).
Culturing of SSC-like cells
We used buffalo Sertoli cells that were treated with 1 μg/μL Mitomycin C (HaiZheng, China) for 2 h as a feeder layer. The SSC-like cells we collected were seeded on the feeder layer and incubated in IMDM culture medium with the known component KSR (Gibco) or the unknown components (FBS), fetuin (Sigma, USA), MEM Non-Essential Amino Acids Solution (10 μL/mL, 100×; Gibco), Lipid mixture 1, Chemically defined (10 μL/mL; Sigma), glial cell-derived neurotrophic factor (GDNF, 20 ng/mL; ProSpec, Israeli), GFRα1 (100 μg/mL; R&D Systems, USA), bFGF (fibroblast growth factor-2, 10 ng/mL, ProSpec), β-Me (2-hydroxy-1-ethanethiol, 0.1 mM, Sangon Biotech, China), B27 (20 μL/mL, Gibco) and LIF2010 (10 U/mL, Millipore, Germany) [14]. The cells were cultured in a 5% CO2 atmosphere at 37°C, and the culture medium was changed every 2 days. The cells were passaged every 4 days.
Propagation of SSC-like cells in vitro
The old culture medium was discarded, and PBS was added to remove cells with poor adherence to the surface. An appropriate amount of 0.05% trypsin (Gibco) was added and allowed to digest the adherent cells for 30 sec, after which the trypsin was neutralized with stem cell culture medium. After gently jetting with a pipette, the surface-adherent cells were collected and resuspended in stem cell culture medium, seeded into a dish coated with gelatine, and incubated for 2 h. Then, the cells in suspension were transferred to a new feeder layer.
Quantitative real-time polymerase chain reaction (PCR)
Total RNA was extracted using a Total RNA Kit I (TaKaRa, China). A PrimeScriptTM RT reagent kit (TaKaRa) with gDNA Eraser was used to synthesize cDNA from the extracted RNA. The genomic DNA was removed by gDNA Eraser for 2 min at 42°C, and the samples were kept at 4°C in a volume of 10 μL. The samples were supplemented with 1 μL of PrimeScriptRT Enzyme Mix I, 1 μL of RT Primer Mix, 4 μL of 5 × PrimeScript Buffer 2 (for Real-Time) and 4 μL of RNase-free ddH2O before being incubated first at 37°C for 15 min and then at 85°C for 5 sec; the samples were then stored at 4°C. Quantitative PCR (q-PCR) was performed on a real-time PCR thermocycler (CFX96 Touch; Bio-Rad, USA). The primer pairs used to amplify messenger RNA (mRNA) are listed in Table 1. The experiment was replicated 3 times. We determined the relative level of expression of each mRNA using the 2−△△Ct (normalized expression ratio) method.
Table 1
Sequences of the primers used in the quantitative real time polymerase chain reaction experiments and the expected product lengths

Induction of SSC-like cells differentiation by retinoic acid (RA) in the culture system with defined materials
The SSC-like cells were cultured in vitro for 2 passages before being gently jetted with a pipette and digested with 0.05% trypsin for 30 sec; the digestion was stopped with IMDM culture medium containing 10% KSR. The collected cells were enriched by the differential plating method and seeded into dishes coated with laminin [28]. In the experimental group, the stem cell culture medium was supplemented with 16 μM RA, while control group cells were cultured in stem cell culture medium. The in vitro induction of SSC-like cell differentiation by RA was continued for 8 days [29]. The cells induced by RA were identified by rabbit anti-human synaptonemal complex protein 3 (SCP3, 1:100; Abcam).
Statistical analysis
All statistical analyses were performed using GraphPad Prism5 (GraphPad Inc., USA). Statistical analyses were performed via Student's t-test or one-way analysis of variance. The results are presented as the mean ± the standard error of the mean. Differences were considered significant when the p value was less than 0.05.
RESULTS
Identification of gene expression in the process of enrichment
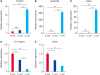
During the enrichment process, we harvested 3 types of cells, G-cells, C-cells and L-cells. As determined by quantitative real-time PCR, the expression levels of the pluripotency marker POU5F1, the marker of inchoate buffalo SSC-like cells NANOS2 (nanos C2HC-type zinc finger 2) and the germ cell marker DDX4 were higher in L-cells than in G-cells and C-cells (p < 0.01, Fig. 1A–C). Furthermore, the expression of β1-integrin and GDNF in G-cells was higher than that in C-cells and L-cells (p < 0.01, Fig. 1D and E).
Fig. 1
Gene expression during the process of purifying G-cells, C-cells and L-cells. Quantitative polymerase chain reaction analysis of the expression of the pluripotency gene POU5F1 (A), germline cell-specific genes NANOS2 (B) and DDX4 (C), and Sertoli cell-specific gene β1-integrin (D) and GDNF (E).
GDNF, glial cell-derived neurotrophic factor; ns, not significant.
*p < 0.05; **0.01 < p < 0.05; ***p < 0.01.
Validation of the SSC-like cells obtained from prepuberal buffalo testicular tissue
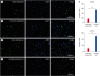
SSCs in adult mice comprise only 0.02% to 0.03% of the total number of cells in the mouse testis [30], which is a very small amount. In our enriched L-cells, the percentages of cells positive for DDX4 and UCHL1 were 55.23% ± 2.23% and 49.67% ± 5.52% of the total cells, respectively (Fig. 2).
Fig. 2
Enrichment of SSC-like cells from testicular cells. Immunocytochemical staining for DDX4 (green) (A) and UCHL1 (green) (C) in testicular cells and DDX4 (green) (B) and UCHL1 (green) (D) in enriched cells. DAPI was used as a counterstain for cell nuclei. Scale bars, 100 μm. (E) Quantitative comparison of the DDX4- and UCHL1-positive cells among testicular and enriched cells. The data are the percentages of the total cell population determined to be DDX4+ and UCHL1+ by immunocytochemical staining.
SSC, spermatogonia stem cell.
***p < 0.01.
In vitro culture of SSC-like cells
In both culture systems, cell clusters were not observed in passage 1 (Supplementary Fig. 1A and D). After 2 days, the cells were passaged at a ratio of 1:2 (passage 2), and no significant target cell proliferation was observed in the culture system with undefined materials (Supplementary Fig. 1B). With continued culture, the target cells gradually disappeared, and the number of fibroblast-like cells increased (Supplementary Fig. 1C). In the culture system with defined materials, the target cells began to proliferate gradually after 2 days, at which point the cells were passaged at a ratio of 1:2 to extend the culture (Supplementary Fig. 1E). As the culture proceeded, the SSC-like cells exhibited a typical string-like morphology as they proliferated (Supplementary Fig. 1F).
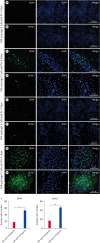
The cells cultured for 2 days and 4 days (passage 2) in both culture systems were fixed at these time points for analysis. In the undefined materials system, the cells cultured for 2 days that were positive for DDX4 and UCHL1 were mainly in the Asingle or Apaired state (Fig. 3A and B). After 4 days of culture, the proportions of DDX4- and UCHL1-positive cells were lower than those after 2 days of culture (Fig. 3C and D). In the culture system with defined materials, positive staining was observed for both DDX4 and UCHL1 in the cells cultured for 2 or 4 days. The cells cultured for 2 days were mainly in the Asingle or Apaired state, and a few cell clones, which are typical proliferative forms of stem cells [14], were also present (Fig. 3E and F). After 4 days of culture, the target cells proliferated, and cell clusters appeared (Fig. 3G and H); however, these clusters were different from the compact and round cell clusters reported in the literature for buffalo [232425]. We recorded the numbers of DDX4- and UCHL1-positive cells (cultured for 2 days at passage 2) that were observable under the microscope in the 2 culture systems, revealing 53.33 ± 4.41 and 46.67 ± 2.61 cells, respectively, in the culture system with defined materials; these numbers were significantly higher than those in the culture system with an unknown composition (18.31 ± 0.88 and 16.67 ± 1.33, respectively) (Fig. 3I).
Fig. 3
Identification of SSC-like cells in the culture systems with undefined materials and defined materials. Buffalo SSC-like cells were cultured on the feeder layers in the systems with undefined materials and defined materials. The SSC-like cells were cultured for 2 days at passage 2 and identified by DDX4 (green) (A) and UCHL1 (green) (B) in the culture system with undefined materials and by DDX4 (green) (E) and UCHL1 (green) (F) in the culture system with defined materials. The SSC-like cells were cultured for 4 days at passage 2 and identified by DDX4 (green) (C) and UCHL1 (green) (D) in the culture system with undefined materials and by DDX4 (green) (G) and UCHL1 (green) (H) in the culture system with defined materials. DAPI was used as a counterstain for cell nuclei. Scale bars in panels (A), (B), (E) and (F), 200 μm. Scale bars in panels (C), (D), (G) and (H), 50 μm. (I) Quantitative comparison of the DDX4- and UCHL1-positive cells from the culture systems with undefined and defined materials. The data were determined by immunocytochemical staining for the number of DDX4- and UCHL1-positive cells observed under the microscope.
SSC, spermatogonia stem cell; KSR, KnockOut Serum Replacement; FBS, foetal bovine serum.
**0.01 < p < 0.05; ***p < 0.01.
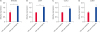
Quantitative real-time PCR analysis of the total RNA in SSC-like cells at passage 2 in both culture systems showed that the gene expression levels of NANOS2, DDX4 and UCHL1 in the culture system with defined materials were significantly higher than those in the culture system with undefined materials (Fig. 4A–C). The expression of cyclin E1 (CCNE1), a gene involved in cell proliferation, was also higher in the system with defined materials than in the undefined materials culture system (Fig. 4D).
Fig. 4
Quantitative PCR analysis of cultured buffalo SSC-like cells at passage 2 in the culture systems with undefined and defined materials. Histograms showing the results of quantitative PCR analysis of the inchoate buffalo SSC-like cell marker NANOS2 (A), germline-related marker DDX4 (B), buffalo SSC-like cell marker UCHL1 (C) and proliferation marker CCNE1 (D) in buffalo SSC-like cells at passage 2 in the culture systems with undefined and defined materials.
PCR, polymerase chain reaction; SSC, spermatogonia stem cell; KSR, KnockOut Serum Replacement; FBS, foetal bovine serum.
*p < 0.05; **0.01 < p < 0.05.
SSC-like cells at passage 3 grown in the culture system with defined materials still proliferated in a serial proliferation mode for 2 days in vitro and exhibited positivity for DDX4 and UCHL1 (Supplementary Fig. 2A and B). When the cells were cultured for 4 days in vitro, the proportions of DDX4- and UCHL1-positive cells were lower than those at 2 days (Supplementary Fig. 2C and D). The immunofluorescence staining results showed that buffalo SSC-like cells could be maintained for approximately 10 days in the culture system with defined materials but for only approximately 6 days in the culture system with undefined materials in vitro.
In vitro induction by RA in the culture system with defined materials
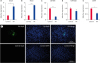
The expression levels of the meiosis differentiation genes stimulated by RA gene 8 (Stra8) (p < 0.05) and meiotic recombination protein REC8 (Rec8) (p < 0.01) were significantly increased. In contrast, the expression levels of the stem cell marker gene ZBTB16 (p < 0.01) and the germ cell marker gene DDX4 (p < 0.05) were significantly decreased. The expression of NANOS2 was also decreased following induction (Fig. 5A–E). In addition, immunofluorescence staining showed that the induced cells were positive for SCP3 (Fig. 5F).
Fig. 5
Quantitative PCR and immunofluorescence analyses of the cultured buffalo SSC-like cells following induction by RA exposure for 8 days in vitro. Histograms showing the quantitative PCR data for the differentiation genes Stra8 (A) and Rec8 (B), SSC-specific markers ZBTB16 (C) and NANOS2 (D), and germline-related marker DDX4 (E). Immunofluorescence staining of the induced cells by SCP3 (F). DAPI was used as a counterstain for cell nuclei. Scale bars, 100 μm.
PCR, polymerase chain reaction; SSC, spermatogonia stem cell; RA, retinoic acid.
*p < 0.05; **0.01 < p < 0.05; ***p < 0.01.
DISCUSSION
Currently, the buffalo SSC culture systems are diversified, and no studies have shown that one system best supports the in vitro culture of buffalo SSCs. The purpose of this study was to compare the effects of culture systems with undefined and defined materials on the in vitro culture of buffalo SSC-like cells to provide a clearer research platform for exploring the mechanism of buffalo SSCs.
Methods for the separation of SSCs from single-cell suspensions mainly include differential plating, magnetic-activated cell sorting (MACS), fluorescence-activated cell sorting (FACS) and Percoll density gradients. The differential plating method was designed based on the differential adhesion abilities of somatic cells and SSCs on different extracellular matrices. SSCs were shown to be prone to adherent growth on laminin, providing a new method for the separation of SSCs [313233]. The differential plating method costs less than MACS and FACS and causes less mechanical damage than Percoll gradient separation [30], leading to better protection of the original pluripotency markers on the cell surface. These benefits of the differential plating method make it conducive to cell culture. In the present study, we enriched the DDX4- and UCHL1-positive cells by the differential plating method, reaching final purities of 55.23% ± 2.23% and 49.67% ± 5.52% (Fig. 2), respectively, which basically met the material requirements for this study.
The molecular markers expressed by different types of cells enriched by the differential planting method were significantly different. The gene NANOS2 is vital for suppressing meiosis [27], while POU5F1, a molecular marker of cellular pluripotency, is expressed in a small subpopulation of SSC-like cells [34], and β1-integrin is secreted by Sertoli cells [35]. Our results showed that the pluripotency and reproductive genes POU5F1, NANOS2 and DDX4 were predominantly expressed in L-cells, whereas β-integrin and GDNF were mainly expressed in G-cells [33] (Fig. 1), as previously described. These results once again demonstrated that we enriched enough buffalo SSC-like cells by the differential plating method.
The growth statuses of the SSC-like cells in the 2 culture systems were different throughout the in vitro culture period. The proliferation of SSC-like cells was very slow in both culture systems during the first cell passage (Supplementary Fig. 1A and D). This phenomenon might have been due to the cells remaining in the recovery phase after thawing. In passage 2 (the cells had survived for 10 days in vitro), the cells cultured in the 2 culture systems were different, with the cell proliferation status, cell number and expression levels of relevant molecular markers in buffalo being better in the culture system with defined materials than in the culture system with undefined materials (Fig. 3). This result was consistent with related reports showing that a low-concentration serum or culture system with defined materials was more suitable for the proliferation of undifferentiated SSC-like cells in vitro [6161925262836]. These results indicated that the unknown components of FBS may be detrimental to the culture of buffalo SSCs in vitro. Compared with FBS, KSR could enhance the proliferation of testicular cells in rats [17]; When FBS was replaced with KSR, a significant increase in the colonization of bovine SSCs and pig undifferentiated SSCs was observable in vitro [1937]. The researchers found that FBS did not contain factors inhibiting SSC colonization and simply lacked factors essential for SSC proliferation [19]. KSR could also promote the proliferation of mammalian embryonic stem cells and induce pluripotent stem cells in vitro [16]. These phenomena may be related to the factors in KSR that promote self-renewal and maintain pluripotency [16]. One of the main components of KSR is AlbuMAX, which contains free fatty acids and cholesterol and plays an important role in cell proliferation [3839]. The disappearance of mitosis is a significant marker of male germ cell differentiation, and studies have shown that KSR can effectively block the stagnation of male germ cell mitosis [1640]. These results were consistent with our results.
In this study, buffalo SSC-like cells could be cultured for only 3 passages in vitro in a culture system with defined materials (Supplementary Fig. 2), which indicated that compared with FBS, KSR could better promote the proliferation and self-renewal of buffalo SSC-like cells in vitro; however, this effect was not permanent. Cells cultured in a culture system with defined materials could be induced into a differentiation state by RA (Fig. 5), indicating that these cultured cells still had SSC characteristics.
In conclusion, the system with defined materials was more conducive to the culture of buffalo SSC-like cells than that with undefined materials. Buffalo SSC-like cells cultured in the system with defined materials had better cell and molecular phenotypes than those cultured in the system with undefined materials. These results could provide a clearer research platform for studying the mechanism underlying buffalo SSC self-renewal and lay a foundation for the future study of more specific systems for culturing buffalo SSCs.
 XML Download
XML Download