

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Phosphoinositide 3-kinases (PI3Ks) are known to play critical roles in inflammation and cancer, promoting cell polarization, migration, adhesion and invasion.1 In mammals, the class I-PI3K isoforms exist in all cell types in which expression of the p110α and p110β isoforms are ubiquitous, while expression of p110γ and p110δ is restricted in some cells such as leukocytes.2 The expression profile of PI3K-δ restricted in white blood cells/immune cells, including lymphocytes and neutrophils, makes it an attractive drug target for various inflammatory or hematological disorders such as rheumatoid arthritis, asthma and chronic lymphocytic leukemia.345
House dust mites (HDMs) are one of the most common aeroallergens worldwide.6 HDM allergic responses are considered to result not only from an exacerbated Th2-biased adaptive immune response but also to be heavily influenced by direct activation of innate immune cells. Consequently, these are associated with the heterogeneity of allergic asthma and its different therapeutic responses. This direct activation is induced by both allergens themselves and danger signals in the allergen sources or the environment.7 The innate immune system senses various allergens including dust mites and molds via pattern recognition receptors (PRRs) such as Toll-like receptor (TLR) and C-type lectin receptor and nucleotide-binding oligomerization domain-like receptors (NLRs). NLRs are divided into 3 subfamilies according to their phylogenetic relationships: NODs, NLRPs and IPAF. Several members of the cytosolic NLRP family act as central components of multiprotein inflammasome complexes which control the production of interleukin (IL)-1β and IL-18.8 Among them, activation of NLRP3 inflammasome is critical for the induction of allergic airway inflammation even steroid-resistant allergic asthma.91011 NLRP3 inflammasome is activated by a variety of signals including pathogen-associated molecular pattern molecules (PAMPs), damage-associated molecular pattern molecules (DAMPs) and bacterial toxins.121314 During activation of innate immune cells by HDM allergens possessing protease allergen and several PAMPs/DAMPs, the first step is PRR/protease allergen receptor activation. Afterward, intricate intracellular signaling activates the NLRP3 inflammasome assembly, which results in the production of inflammatory cytokines and chemokines.15
In addition, a well-known PRR, TLR-mediated intracellular signaling is initiated by its interaction with myeloid differentiation primary response protein 88 (MyD88). MyD88 recruits IL-1 receptor-associated kinase and tumor necrosis factor (TNF) receptor-associated factor to the phosphorylate PI3K/Akt pathway. Subsequently, it induces nuclear factor-κB (NF-κB) nuclear translocation which is closely associated with the gene expression of the NLRP3 inflammasome and pro-inflammatory cytokines.1617 Based on these observations, we have found that HDM-induced allergic responses are associated with the activation of various signaling pathways. However, to date, little information is available on the precise role of PI3K-δ isoform in the assembly of NLRP3 inflammasome or TLR4 pathway in allergic responses.
Therefore, in this study, we aimed to evaluate the role of PI3K-δ in the induction and maintenance of HDM-induced allergic responses in the airway, focusing on the activation of the NLRP3 inflammasome. We used a murine model of HDM-induced asthma in wild type (WT) and PI3K-δ knock-out (KO) mice and also performed an in vitro study using primary cultured murine tracheal epithelial cells and human airway epithelial cells. We present evidence that HDM extract activates the NLRP3 inflammasome via the PI3K-δ signaling pathway linked to nuclear translocation of NF-κB and mitochondrial reactive oxygen species (ROS) and that PI3K-δ can induce the production of epithelial cell-derived cytokines (thymic stromal lymphopoietin [TSLP], IL-25 and IL-33) independently of NLRP3 inflammasome activation in HDM-exposed lung including airway epithelial cells.
MATERIALS AND METHODS
Animals and experiment protocol
Female C57BL/6 mice at 8–10 weeks of age and free of murine-specific pathogens were obtained from the Orient Bio Inc. (Seongnam, Korea). In addition, Cas9 RNA-guided endonuclease (RGEN) PI3K-δ KO mice (Macrogen, Inc., Seoul, Korea) were used, and they were interbred and maintained up to 8–10 weeks of age in pathogen-free condition at Macrogen, Inc. They were housed throughout the experiments in a laminar flow cabinet and maintained on standard laboratory chow ad libitum. All experimental animals used in this study were under a protocol approved by the Institutional Animal Care and Use Committee of Chonbuk National University (CBU 2014-00054). Each animal experiment was performed using 6 mice per group. When using Cas9 RGEN PI3K-δ KO mice, 4 mice were allocated to each experimental group. For the HDM-induced asthma model, mice were sensitized intratracheally with HDM extract (100 μg, Dermatophagoides pteronyssinus; GREER laboratories, Lenoir, NC, USA) on days 1 and 7. On day 14 after the initial sensitization, the mice were challenged intratracheally with 100 μg of HDM extract in saline (or with saline as a control). Bronchoalveolar lavage (BAL) was performed 48 hours after the challenge as previously described.11
Administration of a PI3K-δ inhibitor, IC87114, and an NLRP3 inhibitor, MCC950
IC87114 (0.1 or 1.0 mg/kg body weight/day; Calbiochem, San Diego, CA, USA) or vehicle control (0.05% dimethyl sulfoxide) diluted with 0.9% NaCl was administered in a volume of 50 μL by intratracheal instillation 2 times to each of the treated animals, once at 24 hours before the challenge with HDM and the second time at 24 hours after the challenge. MCC950 (50 mg/kg; Cayman Chemical, Ann Arbor, MI, USA) or vehicle control was administered intraperitoneally once to each mouse at 1 hour before the challenge with HDM.
Histology
Forty-eight hours after the HDM challenge, mice were euthanized. The lung and trachea of each mouse were filled with 10% (volume/volume) neutral buffered formalin intratracheally and were then removed. For fixation, neutral buffered formalin was also used. Specimens were dehydrated and embedded in paraffin. For histological examination, 4-μm sections of fixed and embedded tissues were cut on a Leica model 2165 rotary microtome (Leica Microsystems Nussloch GmbH, Wetzlar, Germany), placed on glass slides, deparaffinized, and stained sequentially with haematoxylin and eosin (Richard-Allan Scientific, Kalamazoo, MI, USA) or periodic acid-Schiff (PAS). Stained slides were analyzed using a light microscope (Axio Imager M1; Karl Zeiss, Goettingen, Germany) under identical conditions including magnification (× 20), gain, camera position and background illumination.18
Quantitation of airway mucus expression
To quantitate the level of mucus expression in the airway, the numbers of PAS-positive and PAS-negative epithelial cells in individual bronchioles were counted as previously described.18
Determination of airway responsiveness to methacholine
Forty-eight hours after the HDM challenge, invasive measurement of airway responsiveness was performed in this study as described elsewhere.19 Anesthesia was achieved through intraperitoneal injection of 45 mg/kg body weight of sodium pentobarbital. The trachea was then exposed through a mid-cervical incision, tracheostomized, and an 18-gauge metal needle was inserted. Mice were connected to a computer-controlled small animal ventilator (flexiVent; SCIREQ, Montreal, Canada). The mouse was quasi-sinusoidally ventilated with the nominal tidal volume of 10 mL/kg body weight at a frequency of 150 breaths/minute and a positive end-expiratory pressure of 2 cm H2O to achieve a mean lung volume close to that during spontaneous breathing. The data was achieved by connecting the expiratory port of the ventilator to the water column. Methacholine aerosol was generated with an in-line nebulizer and administered directly through the ventilator. To determine the differences in airway response to methacholine, each mouse was challenged with methacholine aerosol in increasing concentrations (5.0 to 50 mg/mL in saline). After each methacholine challenge, the data of calculated respiratory system resistance (Rrs) was continuously collected. Changes in Rrs (cmH2O·s/mL) by the methacholine challenge were presented to determine the airway hyperresponsiveness in HDM-inhaled mice.
Western blot analysis
Protein expression levels were analyzed by Western blot analysis as previously described with some modifications.11
Cytosolic and nuclear protein extraction
Cytosolic and nuclear extractions were performed as previously described.11
Measurement of cytokines in BAL fluids or the supernatant of primary cultured cells
The levels of mature IL-1β, cleaved caspase-1, IL-4, IL-5, IL-13, IL-17, and TNF-α in supernatants of cultured cells or BAL fluids were quantified by enzyme immunoassay.
Visualization of mitochondrial ROS using a confocal microscope
BAL cells or primary cultured tracheal epithelial cells were attached to cover-glasses placed on culture dishes for 1 hour and then stained with MitoTracker Red CM-H2Xros (Invitrogen, Carlsbad, CA, USA) in the dark at room temperature. After 30 minutes, cells were washed with phosphate-buffered saline (PBS), and the nuclei were stained using 4′,6-diamidino-2-phenylindole [DAPI] (Invitrogen). The cover-glass having stained cells was mounted on a slide using the fluorescent mounting medium (Golden Bridge International, Inc., Mukilteo, WA, USA) and then visualized using a confocal microscope (Zeiss LSM 510 Meta; Karl Zeiss) equipped with a C-Apochromat 63×/1.20W Korr UV-VIS-IR M27 water immersion objective. Phase contrast microscopy of each group was used for the morphological analysis of cells.
Cell culture and treatment
For the primary culture of tracheal epithelial cells, murine tracheal epithelial cells were isolated from saline- or HDM-instilled mice under sterile conditions as previously described.20 The epithelial cells were seeded onto 60-mm collagen-coated dishes for submerged culture. The growth medium Dulbecco's Modified Eagle Medium (Invitrogen) containing 10% fetal bovine serum, penicillin, streptomycin and amphotericin B was supplemented with insulin, transferrin, hydrocortisone, phosphoethanolamine, cholera toxin, ethanolamine, bovine pituitary extract and bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO, USA). Cells were maintained in a humidified 5% CO2 incubator at 37°C until they adhered. To treat with IC87114 (Calbiochem) or MCC950 (Cayman Chemical), cells were seeded in culture dishes and grown until 70% confluence. The medium was then replaced with a new one containing vehicle, IC87114 (10 μmol/L) or MCC950 (50 nmol/L), and incubated for 2 hours at 37°C.2122
Normal human bronchial epithelial (NHBE) cells were purchased from Lonza (Walkersville, MD, USA). The cells were cultured in bronchial epithelial basal medium (Lonza) supplemented with bovine pituitary extract, gentamicin, amphotericin B, hydrocortisone, epidermal growth factor, epinephrine, insulin, triiodothyronine, transferrin and retinoic acid in a humidified incubator of 5% CO2 atmosphere at 37°C. The cells were seeded in culture dishes and grown until 70% confluence. They were stimulated with 100 μg/mL lipopolysaccharide (LPS) or Der p1 (Indoor Biotechnologies, Cardiff, UK) and 50 ng/mL TNF-α. For the treatment with a TLR4 inhibitor, TAK-242 (1 μmol/L; Calbiochem), and they were pre-incubated in a basal medium containing vehicle (PBS) or TAK-242 at 1 hour before LPS stimulation. Cells were harvested and extracted.
Measurement of phosphatidylinositol (3,4,5)-trisphosphate (PIP3) in lung tissues
A PIP3 competition enzyme immunoassay quantified the amount of PIP3 produced in lung tissues as previously described.4
Immunofluorescence staining for NLRP3, apoptosis-associated speck-like protein containing CARD (ASC), and cleaved caspase-1 in BAL cells and primary cultured tracheal epithelial cells
Primary cultured tracheal epithelial cells or BAL cells were fixed with ice-cold methanol, permeabilized in PBS containing 0.25% Triton X-100 for 10 minutes at room temperature and washed 3 times with PBS. Subsequently, after antigen retrieval for 15 minutes at 37°C in proteinase K (Dako, Glostrup, Denmark), nonspecific bindings were blocked with 1% BSA in PBS containing 0.05% Tween 20 (0.05% PBS-T) for 1 hour. Specimens were then incubated in a humidified chamber for 2 hours at room temperature with an anti-NLRP3 antibody (Adipogen, San Diego, CA, USA), anti-ASC antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) or anti-cleaved caspase-1 antibody (Santa Cruz Biotechnology). For the detection of binding antibody to NLRP3, Alexa Fluor 488 (green) labeled donkey anti-goat IgG (Invitrogen) or Alexa Fluor 546 (red) conjugated anti-mouse IgG (Invitrogen) in 1% BSA was loaded for 1 hour at room temperature in the dark. For the detection of binding antibody to ASC or cleaved caspase-1, Alexa Fluor 488 (green)-labeled anti-rabbit IgG (Invitrogen) was used according to the same protocol. After the specimens were washed, nuclei were stained using DAPI. Stained cells were visualized using a confocal microscope as described above.
Transfection with siRNA in vitro and in vivo
In vitro RNA interference for PI3K-δ and NLRP3 was performed with Stealth RNA interference (Invitrogen) and ON-TARGETplus siRNA (Dharmacon, Lafayette, CO, USA), respectively. For the PI3K-δ knockdown cells, we transfected primary cultured tracheal epithelial cells in the third passage with siRNAs in 6-well plates, but not coated with collagen. Stealth siRNA targeting PI3K-δ or control scrambled siRNA was transfected to the cells grown until 30%–50% confluence. After the transfections, the cells were incubated for 24 hours and then harvested. For transfections, siRNA duplexes were incubated with Lipofectamine RNAiMAX (Invitrogen) according to the manufacturer's instruction. The sequence of Stealth siRNA targeting mouse PI3K-δ was 5′-CAGAUGAGAAGGGAGAGCUGCUGAA-3′ (sense). In the case of NLRP3 interference in NHBE cells, the cells were transfected using ON-TARGETplus siRNA against NLRP3 SMART pool or ON-TARGETplus non-targeting siRNA at a final concentration of 10 nM. The transfection agent was DharmaFECT 4 from ThermoScientific Inc. (Waltham, MA, USA), and Opti-MEM I reduced serum medium (Invitrogen) was used to dilute siRNA and transfection agent according to the manufacturer's protocol. siRNAs (10 nM) were transfected 24 hours before the stimulation of NHBE cells with LPS or Der-p1. As for in vivo experiments of RNA interference, Accell SMART pool siRNA targeting PI3K-δ or control siRNA (5 mg/kg; Dharmacon) was delivered to the murine lung intratracheally once at 24 hours before the challenge with HDM.
Statistical analysis
We used the Shapiro-Wilk test as a test of normality for all data. Normally distributed data was analyzed by using 1-way analysis of variance (ANOVA) followed by Scheffe's test, and statistical comparisons between 2 groups were made by using Student's unpaired t test. Non-normally distributed data was analyzed by using the Kruskal-Wallis ANOVA test followed by the Mann Whitney U test with Bonferroni correction of α = 0.05/κ (κ = number of groups) which was used to reduce the likelihood of false positives due to multiple group comparisons. Statistical comparisons between 2 groups of non-normally distributed data were made by using the Mann Whitney U test. SPSS statistical software (version 19.0; SPSS, Chicago, IL, USA) was used. Data is expressed as mean ± standard error of the mean (SEM). A value of P < 0.05 was considered statistically significant.
RESULTS
Genetic KO and pharmacological blockade of PI3K-δ isoform inhibit HDM-induced PI3K-δ/Akt, NF-κB and TLR4 signaling activation
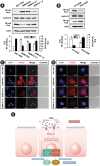
The phosphorylation of Akt and the level of PIP3 in the lung tissues of HDM-instilled mice which indicate the enzyme activity of PI3K were significantly increased compared to those in control mice (Fig. 1A-C). The administration of IC87114 significantly reduced the increased levels of p-Akt and PIP3 in the lung tissues of HDM-instilled mice. As expected, a significant reduction in PI3K-δ isoform expression and the level of p-Akt in lung tissues from HDM-instilled PI3K-δ KO mice were observed compared to those of HDM-instilled WT mice (Fig. 1D-F). In addition, the expression of PI3K-δ mRNA in primary cultured tracheal epithelial cells from HDM-instilled mice was increased by approximately 2-fold compared to that from control mice (Fig. 1G). The transfection of PI3K-δ siRNA reduced PI3K-δ mRNA expression in primary cultured tracheal epithelial cells from HDM-instilled mice up to the control level, whereas control siRNA did not change HDM-induced increase in PI3K-δ mRNA. LPS is one of the macromolecules contained in dust mite extracts. Thus, to evaluate the effect of HDM exposure on airway epithelial cells, human airway epithelial cells were stimulated with LPS. LPS-stimulated airway epithelial cells showed increased levels in p110δ and p85 protein in a time-dependent manner (Supplementary Fig. S1A). The LPS-induced increase of these proteins was significantly reduced by pretreatment with IC87114 (Supplementary Fig. S1B and C). Moreover, LPS-stimulated NHBE cells showed increased levels in p110δ and nuclear NF-κB compared to the levels of unstimulated NHBE cells. Interestingly, the increases of p110δ and nuclear NF-κB protein level in LPS-stimulated NHBE cells were also significantly reduced by pretreatment with a TLR4 inhibitor, TAK-242 (Fig. 1H-J). Conversely, the LPS-induced increases of TLR4 and KC protein expression in NHBE cells (Fig. 1K and L) and the HDM-induced increases in expression of TLR4 in lung tissues were significantly reduced by the administration of IC87114 (Fig. 1M). This data supports that HDM allergen activates the interaction between PI3K-δ and the TLR4 pathway and it can regulate NF-κB activation in the lung, specifically airway epithelial cells.
Fig. 1
Expression of PI3K-δ isoform, TLR4, phosphorylation of Akt and the levels of PIP3 were evaluated in HDM-instilled WT, PI3K-δ KO mice or airway epithelial cells. (A) Representative Western blots of p-Akt in lung tissues from SAL + VEH, HDM + VEH, HDM + IC 0.1 and HDM + IC 1.0 and (B) the densitometric analysis. (C) Enzyme immunoassay of PIP3 in lung tissue extracts. (D) Western blotting of p110δ protein in lung tissues from WT + HDM and δPKO + HDM. (E, F) Representative Western blots of p-Akt in lung tissues and the densitometric analysis. (G) Real-time reverse transcription polymerase chain reaction data showing the levels of PI3K-δ mRNA expression in the primary cultured tracheal epithelial cells. (H) Representative Western blots of p110δ in NHBE cells pre-treated with TAK-242 or vehicle and the densitometric analysis. (I, J) Representative Western blots of NF-κB in NHBE cells pre-treated with TAK-242 or vehicle and the densitometric analysis. (K, L) Representative Western blots of TLR4 (K) and KC (L) in NHBE cells pre-treated with IC87114 or vehicle and the densitometric analyses. (M) Representative Western blots of TLR4 in lung tissues from HDM-instilled mice and the densitometric analysis. Bars represent mean ± standard error of the mean from 4 or 6 mice per group. In vitro experimental data was obtained from 3 independent experiments.
PI3K, phosphoinositide 3-kinase; TLR, toll-like receptor; HDM, house dust mite; WT, wild type; KO, knock-out; NHBE, normal human bronchial epithelial; SAL + VEH, saline-instilled mice with administered with vehicle; HDM + VEH, HDM-instilled mice administered with vehicle; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114; WT + HDM, WT mice instilled with HDM; PKO + HDM, PI3K-δ KO mice instilled with HDM; NF-κB, nuclear factor-κB; LPS, lipopolysaccharide; CON siRNA, control scrambled siRNA; R.D., relative density.
*P < 0.05 vs. HDM + VEH, HDM + CON siRNA, LPS + VEH or WT + HDM; †P < 0.05 vs. SAL + VEH or Control.

PI3K-δ isoform increases protein levels of NLRP3, cleaved caspase-1, and mature IL-1β
Protein expression of NLRP3, cleaved caspase-1 and mature IL-1β was greatly increased in lung tissues (Fig. 2A-F), primary cultured tracheal epithelial cells (Fig. 2G-I) and BAL fluids (Fig. 2J-N) from HDM-instilled WT mice compared to the levels in control mice. Treatment with IC87114 or transfection of siRNA targeting PI3K-δ substantially decreased the expression in these proteins in HDM-instilled WT mice. More interestingly, the protein levels of NLRP3, cleaved caspase-1 and mature IL-1β were markedly lower in lung tissues (Fig. 2O-Q) or BAL fluids (Fig. 2R) of HDM-instilled PI3K-δ KO mice than those in WT mice. Supporting PI3K-δ's involvement in NLRP3 activation at airway epithelial cells, mature IL-1β protein levels were increased in cell lysates (Fig. 2S) as well as supernatants (Fig. 2T) of primary cultured tracheal epithelial cells from HDM-instilled mice, and the treatment with IC87114 dramatically reduced the increases although the statistical significance could not be achieved.
Fig. 2
PI3K-δ increases protein levels of NLRP3, cleaved caspase-1 or mature IL-1β in the HDM-exposed lung. The protein levels were evaluated in HDM-instilled WT and PI3K-δ KO mice. Representative Western blots of NLRP3 (A, D, O), cleaved caspase-1 (B, E, P) and mature IL-1β (C, F, Q) in lung tissues and the densitometric analyses. Representative Western blots of NLRP3 in primary cultured tracheal epithelial cells treated with IC87114 and drug vehicle (G) or transfected with δPI3KD siRNA or CON siRNA (H). (I) Representative confocal laser immunofluorescence photomicrography of primary cultured tracheal epithelial cells shows the expression of NLRP3. Blue fluorescent DAPI stain was used for nuclear localization. Bars indicate a scale of 20 μm. Representative Western blots of cleaved caspase-1 (J) and mature IL-1β (K, L, R) in BAL fluids and the densitometric analysis. Enzyme immunoassay of cleaved caspase-1 (M) and mature IL-1β (N) in BAL fluids of HDM-instilled mice. (S) Representative Western blots for mature IL-1β in cell lysates of primary cultured tracheal epithelial cells treated with IC87114 or drug vehicle. Arrow indicates each target band. (T) Enzyme immunoassay of mature IL-1β in the supernatants of primary cultured tracheal epithelial cells from HDM-instilled mice. Bars represent mean ± standard error of the mean from 4 or 6 mice per group. In vitro experimental data was obtained from 3 independent experiments.
PI3K, phosphoinositide 3-kinase; IL, interleukin; HDM, house dust mite; WT, wild type; KO, knock-out; PI3KD siRNA, siRNA targeting PI3K-δ isoform; CON siRNA, control scrambled siRNA; BAL, bronchoalveolar lavage; HDM + VEH, HDM-instilled mice administered with vehicle; WT + HDM, WT mice instilled with HDM; SAL + VEH, saline-instilled mice with administered with vehicle; SAL, saline-instilled mice; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114; PKO + HDM, PI3K-δ KO mice instilled with HDM; R.D., relative density.
*P < 0.05 vs. HDM + VEH, WT + HDM, or HDM + CON siRNA; †P < 0.05 vs. SAL + VEH or SAL + SAL.

IC87114 reduces HDM-induced NLRP3 inflammasome activation
To visualize NLRP3 inflammasome activation in the lungs of HDM-instilled mice, confocal microscopic examination was performed using BAL cells and primary cultured tracheal epithelial cells. Confocal microscopic analysis showed that the immunofluorescence intensities of NLRP3, ASC and cleaved caspase-1 were dramatically increased and the proteins were co-localized in BAL cells (Fig. 3A and B) and primary cultured tracheal epithelial cells (Fig. 3C and D) from HDM-instilled mice. The treatment with IC87114 restored the increased expressions and co-localization of the proteins in the cells. Additionally, analysis of ASC oligomers by immunoblotting in lung tissues, BAL cells and primary cultured tracheal epithelial cells from HDM-instilled mice revealed the presence of ASC oligomerization (Fig. 3E-G). Furthermore, the data on co-immunoprecipitation of NLRP3 with cleaved caspase-1 or ASC showed that NLRP3 protein interacted with cleaved caspase-1 and ASC to form a protein complex in lung tissues of HDM-instilled mice and the changes were restored by the treatment with IC87114 (Fig. 3H and I).
Fig. 3
Role of PI3K-δ in the assembly of NLRP3 inflammasome in the lung of HDM-instilled mice. (A, B) Representative confocal laser immunofluorescence photomicrography shows the expression of NLRP3 (red), ASC (green in A) and cleaved caspase-1 (green in B) in BAL cells from SAL+VEH, HDM+VEH, HDM+IC 0.1 and HDM+IC 1.0. (C, D) Primary cultured tracheal epithelial cells show the immunofluorescence of NLRP3 (red), ASC (green in C) and cleaved caspase-1 (green in D). The cells were obtained from saline-instilled CON and HDM-instilled mice and were treated with IC87114 (10 μmol/L) or drug VEH. The blue fluorescent DAPI stain was used for nuclear localization. Bars indicate a scale of 20 μm. Merge means the merger views and the square inside of merger view indicate the magnified fields on the right panels. Representative Western blots for ASC oligomer, dimer and monomer in lung tissues (E), BAL cells (F), and in primary cultured tracheal epithelial cells (G) from saline- or HDM-instilled mice. Representative Western blots for caspase-1 (H) and ASC (I) immuno-precipitated with NLRP3 in lung tissues from SAL + VEH, HDM + VEH, HDM + IC 0.1, and HDM + IC 1.0. These data is representative of 6 independent experiments. Arrowheads indicate the target bands.
PI3K, phosphoinositide 3-kinase; HDM, house dust mite; BAL, bronchoalveolar lavage; CON, control mice; VEH, vehicle; DAPI, 4′,6-diamidino-2-phenylindole; SAL + VEH, saline-instilled mice with administered with vehicle; HDM + VEH, HDM-instilled mice administered with vehicle; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114.

Der-p1 protein activates PI3K-δ signaling and NLRP3 inflammasome in NHBE cells
Der-p1-stimulated NHBE cells showed increased levels in p110δ, NLRP3, mature IL-1β and cleaved caspase-1 protein compared to those in unstimulated cells (Supplementary Fig. S2). These findings suggest that several components of HDM extract including Der-p1 contribute to activating PI3K-δ signaling and NLRP3 inflammasome in airway epithelial cells.
PI3K-δ isoform induces NF-κB nuclear translocation and mitochondrial ROS generation
The protein levels of nuclear NF-κB p65 were significantly increased in lung tissues from HDM-instilled mice and these increases were dramatically reduced by treatment with IC87114 (Fig. 4A). PI3K-δ KO mice showed failure in nuclear translocation of NF-κB p65 in lung tissues after HDM stimulation compared to HDM-instilled WT mice (Fig. 4B), suggesting that PI3K-δ isoform is linked to NF-κB activation in the lung of HDM-instilled mice. Additionally, the inhibition of the PI3K-δ isoform reduced increases in mitochondrial ROS generation in BAL cells and primary cultured tracheal epithelial cells from HDM-instilled mice (Fig. 4C and D). These findings suggest that HDM allergens activate the NLRP3 inflammasome in the lung, specifically in airway epithelial cells through activation of the TLR4-PI3K-δ axis, leading to nuclear translocation of NF-κB and mitochondrial ROS generation (Fig. 4E).
Fig. 4
Changes in NF-κB activation and mitochondrial ROS generation in HDM-instilled mice. (A) Representative Western blots of NF-κB in lung tissues from SAL + VEH, HDM + VEH, HDM + IC 0.1 and HDM + IC 1.0, and the densitometric analysis. Bars represent mean ± SEM from 6 mice per group. (B) Representative Western blots of NF-κB in lung tissues from WT + HDM and PKO + HDM, and the densitometric analysis. Bars represent mean ± SEM from 4 mice per group. Representative confocal laser immunofluorescence photomicrographs of BAL cells (C) and primary cultured tracheal epithelial cells (D) show the localization of mitochondrial ROS. The blue fluorescent DAPI stain was used for nuclear localization. Bars indicate a scale of 10 μm (C) or 20 μm (D). (E) Schematic diagram of the role of PI3K-δ isoform in the assembly of the NLRP3 inflammasome and the production of epithelial cell-derived cytokines in the bronchial epithelium of HDM-induced allergic asthmatic mice.
NF-κB, nuclear factor-κB; ROS, reactive oxygen species; HDM, house dust mite; SEM, standard error of the mean; BAL, bronchoalveolar lavage; DAPI, 4′,6-diamidino-2-phenylindole; PI3K, phosphoinositide 3-kinase; SAL + VEH, saline-instilled mice with administered with vehicle; HDM + VEH, HDM-instilled mice administered with vehicle; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114; WT + HDM, WT mice instilled with HDM; PKO + HDM, PI3K-δ KO mice instilled with HDM; R.D., relative density; IL, interleukin; PAMP, pathogen-associated molecular pattern molecule.
*P < 0.05 vs. HDM + VEH; †P < 0.05 vs. SAL + VEH; ‡P < 0.05 vs. WT + HDM.
Blockade of PI3K-δ isoform attenuates HDM-induced asthma features
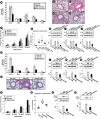
HDM-instilled mice showed typical allergic asthmatic features, the increase in airway inflammatory cells in BAL fluids, infiltration of numerous inflammatory cells around the bronchioles, thickening of airway epithelium in the lung, airway mucus production and the increased airway hyper-responsiveness (Fig. 5A-F). These asthma features were substantially reduced by the administration of IC87114, suggesting that PI3K-δ is an essential contributor to HDM-induced allergic airway inflammation. The HDM-induced increases in IL-4, IL-5, IL-13, KC, IL-17 and TNF-α protein in lung tissues and BAL fluids were also significantly reduced by the administration of IC87114 (Fig. 5G-L, Supplementary Fig. S3). With supporting the data, HDM-instilled PI3K-δ KO mice showed the significant attenuation of asthma manifestations including number of airway inflammatory cells (Fig. 5M), pathological changes (Fig. 5N and O), airway mucus production (Fig. 5N and P), airway hyper-responsiveness (Fig. 5Q and R) and the expressions of Th2 cytokines (Fig. 5S-U) in lung tissues compared to those of HDM-instilled WT mice. Results from experiments using in vivo delivery of siRNA for PI3K-δ isoform to HDM-instilled mice were consistent with these findings as follows; HDM:induced increases in airway inflammatory cells in lung (Fig. 6A), pathological changes (Fig. 6B-E), airway hyper-responsiveness (Fig. 6F and G) and increased expression of Th2 cytokines in lung tissues (Fig. 6H-J) were significantly reduced by the transfection with siRNA targeting PI3K-δ into the mice.
Fig. 5
Effects of IC87114 or genetic KO of PI3K-δ on HDM-induced allergic asthmatic features. (A) The number of inflammatory cells in BAL fluids of saline-instilled or HDM-instilled mice. (B) Representative H&E stained sections (upper panel) and PAS-stained sections (lower panel) of the lung from SAL + VEH, HDM + VEH, HDM + IC 0.1, and HDM + IC 1.0. Bars indicate 50 μm. (C) Inflammation scores of the lung. Total lung inflammation was defined as the average of the peribronchial and perivascular inflammation scores. (D) Quantitation of airway mucus expression. (E, F) Airway responsiveness assessed by invasive (Rrs) measurement in HDM-instilled mice. (G-L) Representative Western blots for IL-4, IL-5, IL-13, KC, IL-17 and TNF-α in lung tissues, and the densitometric analysis. (M) The number of inflammatory cells in BAL fluids of WT + HDM and PKO + HDM. (N) Representative H&E stained sections (upper panel) and PAS-stained sections (lower panel) of the lung from WT + HDM and PKO + HDM. Bars indicate 50 μm. (O) Inflammation scores of the lung. (P) Quantitation of airway mucus expression. (Q, R) Airway responsiveness assessed by invasive (Rrs) measurement. (S-U) Representative Western blots for IL-4, IL-5 and IL-13 in lung tissues, and the densitometric analysis. Bars represent mean ± standard error of the mean from 4 or 6 mice per group.
PI3K, phosphoinositide 3-kinase; HDM, house dust mite; BAL, bronchoalveolar lavage; H&E, haematoxylin and eosin; IL, Interleukin; TNF-α, tumor necrosis factor-α; SAL + VEH, saline-instilled mice with administered with vehicle; HDM + VEH, HDM-instilled mice administered with vehicle; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114; WT + HDM, wild type mice instilled with HDM; PKO + HDM, PI3K-δ knock-out mice instilled with HDM; R.D., relative density; MAC, macrophage; EOS, eosinophil; NEU, neutrophil; LYM, lymphocyte.
*P < 0.05 vs. HDM + VEH or WT + HDM; †P < 0.05 vs. SAL + VEH.
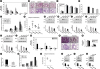
Fig. 6
Changes in asthma features of HDM-instilled mice administered with PI3K-δ targeting siRNA or MCC950. (A, K) The number of inflammatory cells in BAL fluids of HDM-instilled mice. (B-E, O) Representative H&E-stained sections of the lung from HDM-instilled mice. Bars indicate 50 μm. (F, G, P, Q) Airway responsiveness assessed by invasive (Rrs) measurement in HDM-instilled mice. Representative Western blots for IL-4 (H, L), IL-5 (I, M), IL-13 (J, N) and mature IL-1β (R) in lung tissues, and the densitometric analysis. (S) Protein expression of mature IL-1β in BAL fluids quantified by Western blotting. Bars represent mean ± standard error of the mean from 5 or 6 mice per group.
HDM, house dust mite; PI3K, phosphoinositide 3-kinase; BAL, bronchoalveolar lavage; H&E, haematoxylin and eosin; IL, interleukin; CON siRNA, control scrambled siRNA; HDM + VEH, HDM-instilled mice administered with vehicle; SAL, saline-instilled mice; SAL + VEH, saline-instilled mice with administered with vehicle; PI3KD siRNA, siRNA targeting PI3K-δ isoform; R.D., relative density; MAC, macrophage; EOS, eosinophil; NEU, neutrophil; LYM, lymphocyte.
*P < 0.05 vs. HDM + CON siRNA or HDM + VEH; †P < 0.05 vs. SAL + SAL or SAL + VEH.
The inhibition of NLRP3 inflammasome attenuates HDM-induced asthma features
When an NLRP3 inhibitor, MCC950, was administered to HDM-instilled mice, HDM-induced asthma features, including increased airway inflammatory cells (Fig. 6K), expression of Th2 cytokines (Fig. 6L-N), perivascular/peribronchial inflammation (Fig. 6O) and airway hyper-responsiveness (Fig. 6P and Q), were remarkably attenuated with a significant reduction in mature IL-1β in lung tissues (Fig. 6R) and BAL fluids (Fig. 6S).
HDM-induced epithelial cell-derived cytokines are independent of NLRP3 Inflammasome activation
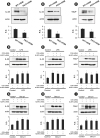
The expression of IL-25, IL-33 and TSLP was dramatically increased in lung tissues (Fig. 7A-C) and primary cultured airway epithelial cells (Fig. 7D-F) from HDM-instilled mice, and these increases were inhibited by the treatment with IC87114. However, these cytokine levels in lung tissues and primary cultured airway epithelial cells were not significantly affected by the treatment with MCC950 (Fig. 7G-L). In addition, PI3K-δ KO mice showed a lower expression level of the cytokines in lung tissues than those of WT mice under HDM exposure (Fig. 8A-C). However, LPS- or Der p1-stimulated NHBE cells (Fig. 8D-I) showed increases in the expression of IL-25, IL-33 and TSLP were persistent even in NLRP3 gene knockdown cells transfected with siRNA. These findings support that epithelial cell-derived cytokines can be affected by the activation of PI3K-δ signaling rather than the activation of NLRP3 inflammasome although NLRP3 inflammasome is associated with HDM-induced asthmatic inflammation.
Fig. 7
Effects of IC87114 or MCC950 on the expression of epithelial cell-derived cytokines in lung tissues and primary cultured tracheal epithelial cells from HDM-instilled mice. Lung tissues were obtained from SAL + VEH, HDM + VEH, HDM + IC 0.1, HDM + IC 1.0 and HDM + MCC950. The cells were obtained from saline-instilled control mice (Control) and HDM-instilled mice, and they were treated with IC87114 (10 μmol/L), MCC950 (50 nmol/L) or drug vehicle. Representative Western blots for IL-25, IL-33 and TSLP in lung tissues (A-C, G-I) and primary cultured tracheal epithelial cells (D-F, J-L), and the densitometric analysis. Bars represent mean ± standard error of the mean from 6 mice per group. In vitro data was obtained from 3 independent experiments.
HDM, house dust mite; SAL + VEH, saline-instilled mice with administered with vehicle; HDM + VEH, HDM-instilled mice administered with vehicle; HDM + IC 0.1, HDM-instilled mice administered with 0.1 mg/kg of IC87114; HDM + IC 1.0, HDM-instilled mice administered with 1.0 mg/kg of IC87114; TSLP, thymic stromal lymphopoietin; IL, interleukin; R.D., relative density.
*P < 0.05 vs. HDM + VEH or Vehicle.; †P < 0.05 vs. SAL+VEH or Control.

Fig. 8
Expression of IL-25, IL-33, and TSLP in lung tissues of HDM-instilled WT and PI3K-δ KO mice (A-C) and in LPS- or Der-p1-stimulated NHBE cells transfected with control or NLRP3 targeting siRNA (D-I). Western blotting of IL-25 (A), IL-33 (B) and TSLP (C) in lung tissues from WT + HDM and δPKO + HDM, and its densitometric analysis. Bars represent mean ± standard error of the mean from 6 mice per group. Representative Western blots for IL-25 (D, G), IL-33 (E, H), and TSLP (F, I) in LPS or Der-p1-stimulated NHBE cells transfected with control or NLRP3 targeting siRNA and the densitometric analyses. Data were from 3 independent experiments.
IL, interleukin; LPS, lipopolysaccharide; HDM, house dust mite; WT, wild type; KO, knock-out; TSLP, thymic stromal lymphopoietin; NHBE, normal human bronchial epithelial; WT + HDM, WT mice instilled with HDM; PI3K, phosphoinositide 3-kinase; PKO + HDM, PI3K-δ KO mice instilled with HDM; CON siRNA, control scrambled siRNA; R.D., relative density.
*P < 0.05 vs. WT + HDM; †P < 0.05 vs. Control.
DISCUSSION
This present study demonstrated that HDM-induced bronchial epithelial activation occurred in both NLRP3 inflammasome-dependent and -independent manners via the PI3K-δ pathway in allergic airway inflammation. In particular, we found that the molecular actions of PI3K-δ signaling played pathophysiological roles in airway epithelial cells as well as immune cells and that PI3K-δ signaling regulates the production of epithelial cell-derived cytokine regardless of NLRP3 inflammasome activation/assembly. Given that PI3K-δ is not the most abundantly expressed PI3K isoform in epithelial cells, our current results are somewhat unexpected and so intriguing.
PI3K-δ is a member of class IA PI3Ks, and it is expressed in inflammatory and immune cell subsets including T cells, B cells and mast cells, which have been shown to play a role in allergic inflammation.23 There is mounting evidence that PI3K-δ plays a pivotal role in the pathogenesis of severe asthma. We have previously reported that a selective PI3K-δ inhibitor, IC87114, improves asthma features and vascular permeability in a murine model of ovalbumin (OVA)-induced allergic asthma.424 Our previous studies have focused on the role and the related mechanisms of PI3K-δ in adaptive immune response, specifically Th2 inflammation424; however, the current study intended to explore the possible role of PI3K-δ isoform as one of the contributors to innate immune response associated with allergic inflammation, specifically NLRP3 inflammasome in bronchial epithelium. While OVA-induced asthmatic inflammation is based on Th2-biased adaptive immune response predominantly, HDM extracts can induce more complex immune responses such as direct activation of innate immune cells. Consistent with the previous observations, current data showed that genetic KO, knockdown with in vivo siRNA transfection, and pharmacological inhibition of PI3K-δ with IC87114 dramatically improved HDM-induced asthmatic manifestations, suggesting that PI3K-δ is an essential contributor to HDM-induced allergic inflammation.
More than 20 HDM allergens belong to protein families with diverse biological functions that contribute to allergenicity.2526 Thousands of proteins and macromolecules contained or produced in a mite, such as LPS, β-glucan and chitin, are PAMPs and may activate innate immunity.2728 Also, DAMPs, such as adenosine triphosphate and uric acid, can be triggered by HDM and are increased in the airways of asthmatic subjects following allergen exposure.2930 LPS is one of the ligands for TLR4 that is usually expressed in epithelial cells and dendritic cells.3132 A previous study has also demonstrated that the expression of TLR4 on airway epithelial cells is up-regulated after challenge with HDM extracts into the airway.33 The activation of TLR4 causes the recruitment of cytosolic adaptor molecules, such as MyD88, resulting in the activation of transcriptional factor NF-κB and mitogen-activated protein kinase pathways.34 Recent studies have also demonstrated that the PI3K-Akt axis activates the downstream of TLRs such as NF-κB and that PI3K-δ is the main class I PI3K isoform recruited to the activated TLR4 complex after stimulation by LPS.353637
Our current data revealed that HDM exposure increases TLR4 expression, Akt phosphorylation and the nuclear translocation of NF-κB in the lung tissues of mice. The inhibition of PI3K-δ suppressed HDM-induced changes in the lung of mice. In addition, the expression of PI3K-δ and nuclear NF-κB proteins was increased in LPS-or Der-p1-stimulated airway epithelial cells and the pretreatment with a TLR4 inhibitor, TAK-242, blocked the increased expression substantially. Moreover, LPS-induced increased KC production in airway epithelial cells was inhibited with the pretreatment with IC87114. These findings suggest that PI3K-δ isoform is one possible downstream pathway of TLR4 stimulation. Altogether, HDM extract-induced PI3K-δ activation is regulated by the TLR4 signaling pathway, which can subsequently activate intracellular signaling molecules such as NF-κB linked to pro-inflammatory mediators in the lung.
Recently, NLRP3 inflammasome has been shown to play a pivotal role in the pathogenesis of HDM-induced atopic dermatitis, in which HDM extracts stimulate assembly of inflammasome in the perinuclear region of keratinocytes.38 NF-κB is involved in the first priming step of activation of inflammasome.153940 In addition, ROS have also been directly implicated as second messengers in the activation of the NLRP3 inflammasome in response to a variety of cellular stress.121314 Moreover, recent studies have revealed that mitochondria are the principal source of ROS required for the assembly of inflammasome.811 Treatment of macrophages with Mito-TEMPO, a scavenger of mitochondrial ROS, results in inhibition of NLRP3 inflammasome activation.8 Furthermore, another mitochondrial ROS scavenger, NecroX, attenuates steroid-resistant asthma symptoms successfully through inhibiting of NLRP3 inflammasome.11 Our current data revealed that inhibition of PI3K-δ blocked nuclear translocation of NF-κB, thereby reducing the production of IL-4, IL-5, IL-13 and IL-17 in the lung of HDM-instilled mice. We also found that mitochondrial ROS generation was significantly increased in BAL cells and primary cultured tracheal epithelial cells isolated from HDM-instilled mice, and that IC87114 markedly reduced the increases of mitochondrial ROS generation. In addition, we found that MCC950 significantly decreased HDM-induced allergic asthma features in mice. As for the direct role of PI3K-δ in NLRP3 inflammasome assembly, our current data showed that blockade of PI3K-δ substantially decreased the HDM-induced protein levels of NLRP3, cleaved caspase-1 and mature IL-1β in the lung including epithelial cells and even its supernatants. Furthermore, NLRP3, ASC and caspase-1 were dramatically increased, co-localized and united into the inflammasome complex in BAL cells and primary cultured tracheal epithelial cells from HDM-instilled mice, and the treatment with IC87114 restored these changes. These results suggest that PI3K-δ signaling regulates the assembly of the NLRP3 inflammasome in lung including epithelial cells. Altogether, these findings indicate that PI3K-δ signaling is implicated in the assembly of the NLRP3 inflammasome in the HDM-exposed airway cells via NF-κB activation and mitochondrial ROS generation.
In addition to the traditional barrier function, airway epithelium plays immune/inflammatory roles in asthma pathogenesis.41 Microbial ligands activate TLRs on airway epithelial cells, thereby generating a significant amount of TSLP that enhances Th2 polarization as well as enables mast cells to secrete selective cytokines.4243 These findings seem to be one of the mechanisms explaining how airway epithelial cells can provide a microenvironment to sustain ongoing Th2 inflammation and how microbial stimuli contribute to aggravating asthma. In our study, the results revealed that the expression levels of PI3K-δ mRNA or proteins, p110δ and p85 were substantially increased in primary cultured airway epithelial cells from HDM-instilled mice and LPS-stimulated human airway epithelial cells. The increases were significantly reduced by the pretreatment with siRNA targeting PI3K-δ, IC87114 or TAK-242 in vitro.
Moreover, blockade of PI3K-δ substantially reduced the expression of epithelial cell-derived cytokines (i.e., IL-25, IL-33 and TSLP) in lung tissues and epithelial cells from HDM-instilled mice. Given the restricted expression of PI3K-δ in leukocytes, these results are very worthy of note. Based on our findings regarding NLRP3 assembly in primary cultured epithelial cells, we carefully propose that airway epithelial cells can act as an immune cell through a novel role of PI3K-δ isoform as a bridge between 2 PRRs, TLR4 and NLRP3, resulting in complicating allergic responses. Another interesting finding is that MCC950 and NLRP3 knockdown procedure using siRNA transfection failed to inhibit HDM-induced epithelial cell cytokine production, suggesting that epithelial cell-derived cytokines do not depend on the activation of the NLRP3 inflammasome and that PI3K-δ is more potent controller than NLRP3 inflammasome for allergic airway inflammation, especially epithelial cell-derived Th2 responses.
In summary, HDM allergens activate NLRP3 inflammasome in the lung through TLR4-PI3K-δ axis activation linked to nuclear translocation of NF-κB and mitochondrial ROS generation. At the same time, PI3K-δ signaling can induce epithelial cell activation in an NLRP3 inflammasome-independent manner. The novel roles of PI3K-δ in the assembly of NLRP3 inflammasome and the production of epithelial cell-derived cytokines highlight the therapeutic potential of PI3K-δ targeting agents as well as the pathophysiological role of airway epithelial cells as an immune responder in allergic inflammation.
 XML Download
XML Download