

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The production of antibodies is an essential arm of the immune response that provides both immediate protection against invading antigens or self-antigens and long-term immunity when reexposed to the same antigens. The antibody-secreting cell compartment consists of both plasmablasts and plasma cells. The plasmablasts are proliferating precursors of plasma cells, whereas plasma cells are end-differentiated cells.12 Based on their lifespan, plasma cells are generally divided into short- and long-lived populations. The long-lived plasma cells (LLPCs) can survive in bone marrow for decades with durable antibody-producing capacity, even in the absence of antigens after the original antigen encounter, which is important for humoral immunity.34 LLPCs do not proliferate, whereas they express high levels of anti-apoptotic protein B cell lymphoma 2 (BCL2).15
Studies in humans and mice have shown that LLPCs reside in specific survival niches in the bone marrow, tonsils, and spleen,12346 depending on soluble factors in the local milieu and physical interactions with surrounding stroma.789101112 It has been demonstrated that interleukin (IL)-6, IL-5, IL-21, and tumor necrosis factor α are involved in the formation of specific micro niche beneficial to the development of LLPCs in bone marrow.34 The pool of these survival factors for plasma cells in bone marrow is referred to as survival niches. Recently, increasing evidence suggests that LLPCs can also survive in inflammatory tissues, such as the lungs of asthmatic mice and the kidneys of lupus mice,1314 and therefore maintain inflammatory and autoimmunity processes. Nerve growth factor (NGF) and neurotrophin (NT) 3 belong to the NT family and are potent in mediating growth, proliferation, and survival of various cell types including plasma cells and B cells.15161718 NGF and NT3 have been indicated to play a critical role in creating a survival micro niche for plasma cells in inflamed airways by activating of an anti-apoptotic pathway in plasma cells.13 Although the durability of LLPCs is beneficial in the context of immunity to infection and vaccination, LLPCs may exaggerate and prolong the inflammation process in immunoglobulin (Ig)-mediated allergic inflammation and autoimmune diseases.1319 In addition, LLPCs are refractory to glucocorticoid treatment, irradiation, and immunosuppression, making them a potential factor contributing to the relapse of diseases after treatment in patients with allergic and autoimmune diseases.16
Chronic rhinosinusitis with nasal polyps (CRSwNP) is characterized by persistent inflammation of nasal and paranasal sinus mucosa with the formation of nasal polyps.20 According to the extent of eosinophil infiltration in polyps, CRSwNP is subclassified into eosinophilic and non-eosinophilic CRSwNP, especially in Asian patients.21 Local Ig production has been found to be associated with disease pathogenesis and treatment outcome in patients with CRSwNP.2223242526 Local IgA, IgG and IgD levels have been demonstrated to be elevated in both eosinophilic and non-eosinophilic CRSwNP, whereas local IgE has been shown to be upregulated only in eosinophilic CRSwNP.22232425 Previously, we and others have shown enhanced local infiltration of plasma cells in both eosinophilic and non-eosinophilic CRSwNP.21222728 However, whether the plasma cells in nasal polyps contain a long-lived population, and whether the nasal polyp environment provides a specific survival niche for LLPCs remain unknown.
In this study, we identified Ki-67−BCL2+ plasma cells in nasal polyps, which possibly contribute to the long-time Ig production in nasal polyps. Elevated expression of NGF in nasal polyps may provide a micro environment for the development of LLPCs with the expression of tropomyosin receptor kinase (Trk) A, an NGF high-affinity receptor.
MATERIALS AND METHODS
Subjects and sample collection
The study was approved by the Ethics Committee of Tongji Hospital of Tongji Medical College of Huazhong University of Science and Technology (Permit Number 20160301) and conducted with written informed consent from each patient. The diagnosis of CRSwNP was made according to the current European Position Paper on Rhinosinusitis and Nasal Polyps (EPOS2012).20 CRSwNP was defined as eosinophilic when the percentage of tissue eosinophils exceeded 10% of total infiltrating cells, as reported by our previous study.21 This cutoff was calculated as twice the standard deviation of the mean eosinophil percentage in control subjects.21 Patients undergoing septoplasty because of anatomic variation and without other sinonasal diseases were enrolled as control subjects. Asthma was diagnosed based on the Global Initiative for Asthma guidelines.29 The diagnosis of allergic rhinitis was made based on the concordance between a typical history of allergy symptoms and positive atopy test.30 Atopic status was evaluated with a skin prick test using a standard panel of aeroallergens common in our region and/or ImmunoCAP (Thermo Fisher Scientific, Uppsala, Sweden) to detect IgE antibodies against common inhalant allergens. Intranasal steroid sprays and oral glucocorticoids were stopped at least 1 month and 3 months before surgery, respectively. None of the patients had received antileukotrienes or immunotherapy. Subjects who had cystic fibrosis, antrochoanal polyps, fungal sinusitis, gastroesophageal reflux disease, primary ciliary dyskinesia, vasculitis, or an acute upper respiratory tract infection within 1 month of entering the study were excluded from the study. None of the patients had a history of aspirin sensitivity. The demographic and clinical data of the subjects are summarized in Supplementary Table S1.
Nasal polyp tissues from patients with CRSwNP and inferior turbinate mucosal samples from control subjects were harvested during surgery. Due to the limited amount of tissue samples, not all samples were included in every study protocol. Fresh surgical specimens were transported on ice-cold Roswell Park Memorial Institute 1640 medium (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) to the laboratory for further processing within minutes after sampling.
Ex vivo nasal tissue culture
Fresh nasal polyp and inferior turbinate mucosal samples were sectioned into multiple pieces of approximately 2 to 3 mm3. Some tissue sections were prepared for histological study directly. Some tissue sections were subjected to an ex vivo air-liquid interface culture. The remaining sections of tissue samples were preserved at −80°C for RNA extraction.
The ex vivo culture was performed as previously described.31 Briefly, tissue sections were placed on 0.4-µm well inserts (Millipore Corp., Billerica, MA, USA) in 2 mL of Dulbecco modified Eagle medium/F-12 (Gibco, Thermo Fisher Scientific) supplemented with 10% fetal calf serum and penicillin/streptomycin (Guge Biotechnology, Wuhan, China) at 50 μg/mL in 6-well trays. The tissue samples were oriented with the epithelium exposed to the air, forming an air-liquid interface to mimic the in vivo situation, and cultured in a 5% CO2-humidified atmosphere at 37°C. The samples were weighted and 3 tissue sections per well were cultured in duplicate to minimize discrepancies related to variations in sample size and handling. To reduce the passive shedding of Igs previously deposited in tissues, the culture medium was refreshed 1 day after culture. Nasal tissue sections were cultured for 32 days with complete medium refreshment every 2 days. The harvested culture supernatants were centrifuged and preserved at −80°C for further analysis.
Measurement of Ig levels
The levels of Igs in culture supernatants were detected using commercially available enzyme-linked immunosorbent assay (ELISA) kits (Senxiong, Shanghai, China) as previously reported.2425 The concentrations of Igs were normalized by the sample weight. Lower detection limits for the ELISA assays are shown in Supplementary Table S2.
Histologicaly analysis
Paraffin sections (4 µm) were prepared. Hematoxylin and eosin staining, immunohistochemistry, and immunofluorescence staining were performed as previously described.212224 For immunohistochemical staining, antigen retrieval was performed using the Target Retrieval Solution (Dako, Carpinteria, CA, USA). Hydrogen peroxide (3%) and normal serum (Boster Biotechnology, Wuhan, China) were applied to block endogenous peroxidase activity and non-specific binding, respectively. The sections were stained with primary antibodies as listed in Supplementary Table S3. Species and isotype-matched antibodies were used as negative controls. Antigens were detected using the streptavidin-biotin-peroxidase complex method with a histostain-plus kit (Boster Biotechnology) as previously described.22 Color development was achieved with 3′, 3′-diaminobenzidine, which rendered positive cells brown. The intensity of NGF and TrkA positive staining in the epithelial cells was analyzed using Image-Pro® Plus software (Media Cybernetics, Inc., Rockville, MD, USA) and the average integrated optical density (IOD) per stained area (IOD/area) was calculated as previously described.32 Consecutive sections were used to determine the relationship between eosinophilic cationic protein (ECP) positive eosinophils and the expression of NGF.
Double immunofluorescence staining was used to determine the relationship between CD138+ plasma cells as well as BCL2+, Ki67+, and TrkA+ cells, and between tryptase-positive mast cells and NGF-positive cells. The fluorescence-conjugated secondary antibodies used are shown in Supplementary Table S4.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from sinonasal mucosal samples using TRIzol (Invitrogen, Carlsbad, CA, USA) and treated with DNase (TaKaRa Biotechnology, Dalian, China) to remove DNA contamination. One microgram RNA was reverse-transcribed to cDNA with random hexamer primer.22 One microliter of cDNA was used to perform quantitative polymerase chain reaction, which was performed with specific primers (Supplementary Table S5) using a SYBR Premix Ex Taq kit (TaKaRa Biotechnology) as previously mentioned.22 Relative gene expression was calculated using the comparative computed tomography method.2122 Then, β-actin was used as a housekeeping gene for normalization and a no template sample was used as a negative control.
Western blotting
Western blot analysis was performed as previously described with minor modifications.3435 Total protein was extracted from fresh surgical specimens with radio-immunoprecipitation assay lysis buffer containing a 2% cocktail of protease inhibitor (Guge Biotechnogogy). Protein concentrations were measured with a BCA protein assay kit (Guge Biotechnology). Samples containing 40 μg of proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore Corp.). The membranes were blocked with 5% fat-free milk in Tris-buffered saline with Tween-20 (Guge Biotechnology). Primary antibodies as listed in Supplementary Table S3 were incubated with PVDF membranes overnight at 4°C. Then, the membranes were incubated with horseradish peroxidase conjugated secondary antibody (goat anti-rabbit, 1:5,000; Guge Biotechnology) for 1 hour at room temperature. After that, the members were processed using an ECL chemiluminescence reaction kit (Guge Biotechnology), followed by exposure on chemiluminescent film to visualize the proteins. β-actin was used as an internal control to correct for variations in protein loading. Densitometric analysis of the blots was performed using AlphaEase FC software (Alpha Innotech, Silicon Valley, CA, USA). Relative protein expression was quantified and normalized to β-actin expression.
Statistical analysis
Statistical analysis was performed with the SPSS 18.0 software (SPSS Inc., Chicago, IL, USA). For continuous variables, data are presented in dot plots unless stated otherwise. Symbols represent individual samples, horizontal bars represent medians, and error bars show interquartile ranges. When comparisons were made among groups, the Kruskal-Wallis H test was used to assess significant inter-group variability. The Mann-Whitney U 2-tailed test was used for between-group comparison. The significance was accepted at P < 0.05.
RESULTS
Increased accumulation of BCL2+CD138+ plasma cells in nasal polyps
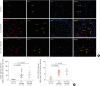
The expression of BCL2, a critical anti-apoptotic molecule, in plasma cells prevents the apoptosis of plasma cells and leads to the development of LLPCs.36 In addition to confirming the increased infiltration of CD138+ plasma cells in both eosinophilic and non-eosinophilic polyps in comparison to control tissues (Fig. 1), we found that the numbers of BCL2+CD138+ plasma cells were increased in both eosinophilic and non-eosinophilic polyps by immunofluorescence staining (Fig. 1).
Fig. 1
Increased accumulation of BCL2+CD138+ plasma cells in nasal polyps. (A) Representative photomicrographs showing BCL2+CD138+ plasma cells in control tissues, and in eosinophilic and non-eosinophilic nasal polyp tissues. Red arrows indicate representative CD138+ plasma cells; green arrows indicate representative BCL2+ cells; yellow arrows indicate representative BCL2+CD138+ plasma cells (original magnification ×400). (B) Quantification of CD138+ total plasma cells and BCL2+CD138+ plasma cells in lamina propria.
BCL2, B cell lymphoma 2; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps.
Sustained Ig production from ex vivo cultured polyp tissues
Our previous study demonstrated higher numbers of IgA+ and IgG+ plasma cells in eosinophilic and non-eosinophilic polyps and higher numbers of IgE+ plasma cells in eosinophilic polyps than those in control tissues.24 No upregulation of IgM+ plasma cells were found in polyps, which was in line with the comparable IgM levels among different groups.24 To explore whether there is a long-lived population of plasma cells in nasal polyps, we used an ex vivo nasal tissue culture model, since the intact tissue structure would provide optimal survival conditions for Ig-secreting cells.23 We chose 32-day as the time period for culture, since the life span of conventional plasma cells is approximately 3 to 5 days, and the life span of LLPCs in the human small intestine is approximately 32 days.3
As shown in Fig. 2, we found that IgG was the most abundant Ig isotype produced in nasal polyps, followed by IgA, IgM, and IgE. Compared to control tissues, the production of IgA and IgG was upregulated in both eosinophilic and non-eosinophilic polyps, and the production of IgE was enhanced only in eosinophilic polyps (Fig. 2). Although the levels of IgA and IgG produced from both eosinophilic and non-eosinophilic polyp tissues, and IgE levels produced from eosinophilic polyps decreased dramatically in the first 8 days during culture, they retained a relatively stable level for the next 20 days (Fig. 2), suggesting the potential presence of LLPCs for the sustained Ig production in polyp tissues. We did not find a significant difference in the secretion of IgM from polyp tissues and control tissues (Fig. 2).
Fig. 2
Ig production from ex vivo cultured sinonasal mucosal samples. Eosinophilic and non-eosinophilic nasal polyps and inferior turbinate mucosal samples were cultured ex vivo for 32 days. Culture supernatants were collected at days 2, 4, 8, 16, and 32, and Ig levels in the culture supernatants were measured by enzyme-linked immunosorbent assay.
Ig, immunoglobulin; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps.

The preservation of Ki-67−BCL2+ plasma cells in nasal polyp tissues after ex vivo culture
To delineate the mechanisms underlying the durable Ig production in nasal polyps, we studied histological changes in nasal polyps after a 32-day culture. We found that the morphology of tissue structure, especially that of epithelium, preserved well after a 32-day ex vivo culture (Supplementary Fig. S1). Although the numbers of plasma cells were decreased significantly in the post-culture polyp tissues compared with those in tissues before culture, there were certain numbers of plasma cells retained in polyp tissues (median, 15 plasma cells/high power field) after the 32-day culture (Fig. 3A). This is in contrast to the findings of other immune cells. Few CD20+ B cells, and CD3+, CD4+ and CD8+ T cells could be detected in polyp tissues after the 32-day culture (Supplementary Fig. S2A). The percentages of reduction of cell numbers of CD20+ B cells, and CD3+, CD4+ and CD8+ T cells after culture were significantly higher than that of plasma cells (Supplementary Fig. S2B).
Fig. 3
BCL2+CD138+ plasma cells are retained in nasal polyps after a 32-day ex vivo culture. (A) Representative photomicrographs showing CD138+ plasma cells in nasal polyp tissues before and after culture and the quantification of CD138+ plasma cells. Arrows indicate representative CD138+ plasma cells. (B) Representative photomicrographs and quantitative analysis of BCL2+CD138+ plasma cells in nasal polyp tissues before and after culture. Red arrows indicate representative CD138+ plasma cells; green arrows indicate representative BCL2+ cells; yellow arrows indicate representative BCL2+CD138+ plasma cells. (C) Representative photomicrographs show that CD138+ plasma cells in nasal polyp tissue do not have Ki-67 expression before and after culture. Red arrows indicate representative CD138+ plasma cells; green arrows indicate representative Ki67+ cells; white arrows indicate representative Ki67−CD138+ plasma cells (original magnification ×400).
BCL2, B cell lymphoma 2.
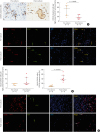
Interestingly, different from the change in total plasma cells, we found that the numbers of BCL2+CD138+ plasma cells were comparable before and after culture, and the percentage of BCL2+CD138+ plasma cells in total plasma cells was increased after the 32-day culture (Fig. 3B). In addition, we found that CD138+ plasma cells in nasal polyp tissues did not express Ki-67 (Fig. 3C), indicating that BCL2+ plasma cells with longer life span in nasal polyps are non-proliferative.
Upregulation of NGF and TrkA in nasal polyps
To determine whether the local environment in nasal polyp tissues provides essential factors for sustaining the long-term survival of plasma cells, we measured the expression of NTs including NGF and NT3, which are involved in creating a survival micro niche for plasma cells in airways.15
We found that the mRNA expression of NGF, brain derived neurotrophic factor (BDNF), and NT3 was upregulated in both eosinophilic and non-eosinophilic polyps compared with control tissues (Fig. 4A). The mRNA expression of TrkA, a high-affinity receptor for NGF, was upregulated in eosinophilic and non-eosinophilic CRSwNP compared with controls (Fig. 4B). However, there was no change in mRNA expression of p75 neurotrophin receptor (P75), a low affinity receptor for NGF, or TrkB, a receptor for BDNF, or TrkC, a receptor for NT3, in eosinophilic and non-eosinophilic polyps compared with control tissues (Fig. 4B). In addition, a strong correlation between NGF mRNA expression level and BCL2+CD138+plasma cell count was discovered in nasal tissues (Fig. 4C). Moreover, we observed that TrkA and NT3 mRNA expression levels were modestly correlated with BCL2+CD138+plasma cell count in nasal tissues (Fig. 4C). No significant correlation between BDNF mRNA expression level and BCL2+CD138+plasma cell count in nasal tissues was found (Fig. 4C). These data suggest that the NGF/TrkA signaling pathway may play a role in plasma cell survival in nasal polyps.
Fig. 4
The mRNA expression of NTs (A) as well as their receptors (B), and correlations between BCL2+CD138+ plasma cell count and the expression level of NTs and their receptors (C) in sinonasal mucosa samples as detected by quantitative real-time polymerase chain reaction.
NT, neurotrophin; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps; NGF, nerve growth factor; BDNF, brain derived neurotrophic factor; Trk, tropomyosin receptor kinase; BCL2, B cell lymphoma 2; P75, p75 neurotrophin receptor.

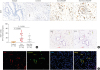
Next, the protein expression of NGF and TrkA in nasal tissues was investigated by immunohistochemistry. We found that NGF was expressed by nasal epithelial cells and inflammatory cells in the lamina propria (Figs. 5 and 6A). A higher staining intensity of NGF in nasal epithelial cells was detected in eosinophilic and non-eosinophilic nasal polyps compared with that in control nasal tissues (Fig. 5). In addition, the numbers of NGF+ cells in the lamina propria were increased in both eosinophilic and non-eosinophilic nasal polyps compared with control tissues (Fig. 6A). ECP+ eosinophils and tryptase+ mast cells had the expression of NGF as detected by immunohistochemical staining of serial tissue sections and double immunofluorescence staining, respectively (Fig. 6B and C). In addition, few CD3+ T cells also expressed NGF (data not shown).
Fig. 5
Increased protein expression of NGF in epithelial cells in eosinophilic and non-eosinophilic nasal polyps. Representative photomicrographs showing NGF expression in nasal epithelial cells and infiltrating cells in control tissues and eosinophilic and non-eosinophilic nasal polyp tissues as detected by immunohistochemistry. Isotype control staining is also shown. Arrows indicate representative NGF+ cells. The expression intensity of NGF in epithelial cells was quantified (original magnification ×400).
NGF, nerve growth factor; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps; IOD, integrated optical density.
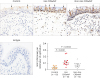
Fig. 6
Increased protein expression of NGF in lamina propria in eosinophilic and non-eosinophilic nasal polyps. (A) Representative photomicrographs showing NGF+ cells in the lamina propria of control tissues, and eosinophilic and non-eosinophilic nasal polyp tissues as detected by immunohistochemistry. Arrows indicate representative NGF+ cells. The numbers of NGF+ cells in the lamina propria were quantified. (B) Representative immunostaining of consecutive tissue sections from a patient with eosinophilic chronic rhinosinusitis with nasal polyps (Eos CRSwNP) showing the expression of NGF by ECP+ eosinophils. Arrows with the same direction indicate the same cells in consecutive serial sections. (C) Representative double immunofluorescence staining of a tissue section from a patient with Eos CRSwNP showing the expression of NGF by tryptase-positive mast cells. Red arrows indicate representative NGF+ cells; green arrows indicate representative tryptase+ cells; yellow arrows indicate representative NGF+tryptase+ cells (original magnification ×400).
NGF, nerve growth factor; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; ECP, eosinophilic cationic protein; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps. HPF, high power field.
TrkA immunoreactivity was mainly found in inflammatory cells in the lamina propria and epithelial cells (Fig. 7A and Supplementary Fig. S3). The number of TrkA+ cells in the lamina propria and the staining intensity of TrkA in nasal epithelial cells were significantly increased in both eosinophilic and non-eosinophilic nasal polyps compared with controls (Fig. 7A and Supplementary Fig. S3). More importantly, TrkA expression was identified on CD138+ plasma cells in the lamina propria of nasal polyps (Fig. 7B).
Fig. 7
Increased protein expression of TrkA in eosinophilic and non-eosinophilic nasal polyps. (A) Representative photomicrographs showing TrkA expression in infiltrating cells in the lamina propria of control tissues, and eosinophilic and non-eosinophilic nasal polyp tissues as detected by immunohistochemistry. Arrows indicate representative TrkA+ cells. The numbers of TrkA+ cells in the lamina propria were quantified. (B) Representative double immunofluorescence staining of a tissue section from a patient with eosinophilic chronic rhinosinusitis with nasal polyps (Eos CRSwNP) showing the expression of TrkA by CD138+ plasma cells. Green arrows indicate representative TrkA+ cells; red arrows indicate representative CD138+ cells; yellow arrows indicate representative TrkA+CD138+ plasma cells (original magnification ×400).
Trk, tropomyosin receptor kinase; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps. HPF, high power field.
To further confirm the protein levels of NGF and TrkA in nasal tissues, Western blot analysis was performed. Consistent with our immunohistochemistry results, we found that NGF and TrkA protein expression was upregulated in eosinophilic and non-eosinophilic nasal polyps compared with control tissues (Fig. 8A-C).
Fig. 8
Increased protein expression of NGF and TrkA in eosinophilic and non-eosinophilic nasal polyps detected by western blot analysis. (A) Representative blots of NGF and TrkA in whole—cell lysates of nasal tissues are shown. (B) NGF relative protein expression was quantified and normalized to β-actin expression. (C) TrkA relative protein expression was quantified and normalized to β-actin.
NGF, nerve growth factor; Trk, tropomyosin receptor kinase; Eos CRSwNP, eosinophilic chronic rhinosinusitis with nasal polyps; Non-Eos CRSwNP, non-eosinophilic chronic rhinosinusitis with nasal polyps.

DISCUSSION
Although the presence of plasma cells in nasal polyps may contribute to local Ig production, whether there are LLPCs in nasal polyps has not been studied previously. BCL2 expression extends the survival of plasma cells by preventing apoptosis.36 Consistent with this notion, we found that BCL2+CD138+ plasma cells in nasal polyps survived after a 32-day ex vivo culture, although the total number of plasma cells reduced significantly. This finding is in line with the reduced but sustained Ig production in nasal polyps after long-time culture without antigen exposure. Previous studies showed that the life span of LLPCs in the human small intestine and tonsils is approximately 32 and 13 days, respectively.23 Therefore, in this study, we chose 32 days as the time period for culture and our data indicate the presence of LLPCs in nasal polyps after a 32-day ex vivo culture. In the present study, we did not find the expression of proliferation marker Ki-67 in the CD138+ plasma cells either before or after culture, suggesting that these LLPCs are non-proliferative cells, which is consistent with previous finding that BCL2 expression inversely correlated with the proliferation rate of malignant plasma cells.3637 Moreover, we discovered that the numbers of BCL2+CD138+ plasma cells were increased in both eosinophilic and non-eosinophilic CRSwNP compared with controls. Collectively, these data suggest the involvement of LLPCs in the sustained Ig production in CRSwNP even without antigen exposure.
The local microenvironment is critical for the generation of LLPCs.239 Previous studies have demonstrated that NTs promote the survival of plasma cells in airways.1318383940 Studies using a murine asthma model have demonstrated that NGF plays a critical role in creating a survival micro niche for Ig secreting plasma cells in inflamed airways by induction of the anti-apoptotic pathway, and that the overexpression of NGF in epithelial cells is accompanied by an increase in CD138+ plasma cells in the perialveolar area.13 Similar to the findings in inflammatory lung tissue,1520 here we suggest a NT driven micro-environment in supporting plasma cells survival in nasal polyps. We found that the expression of NGF was significantly upregulated in both eosinophilic and non-eosinophilic nasal polyps compared with control tissues. Consistent with studies of allergic rhinitis, we found that NGF was mainly expressed by epithelial cells in nasal polyps.40 In our 32-day culture system, the structure of epithelial cells was well preserved, which may persistently supply NGF for LLPC survival in nasal polyps. In addition, eosinophils, mast cells, and few T cells were also found to be the source of NGF in nasal polyps. Importantly, we further demonstrated that CD138+ plasma cells had the expression of TrkA, the high affinity receptor for NGF, in nasal polyps, supporting the involvement of the NGF/TrkA pathway in the development of LLPCs in nasal polyps.
There are several limitations of our current study. First, plasma cells may be locally newly generated as nasal polyps contain B cells.22 However, CD20+ B cells were markedly diminished after the 32-day culture, suggesting that newly generated plasma cells unlikely contribute to the sustained Ig production after long-time culture. Secondly, there are currently no generally accepted markers to define LLPCs. Although BCL2 extends the survival of plasma cells by anti-apoptosis,36 BCL2+CD138+ plasma cells are not considered to be LLPCs. We therefore performed long-time culture to prove the presence of LLPCs in nasal polyps. We found that in line with the sustained Ig production, BCL2+CD138+ plasma cells remained after culture. Our study indicates that BCL2+CD138+ plasma cells may reflect the presence of LLPCs in nasal polyps. Thirdly, a number of factors have been reported to modulate the generation of LLPCs. In the present study, we concentrated on NTs, which have been reported to regulate the survival of plasma cells in the lungs. Whether there are other factors involved in LLPC generation in nasal polyps requires further studies.
In conclusion, this study demonstrated that human nasal polyps harbor a nonproliferative Ig secreting plasma cell population surviving for more than 4 weeks, that is, LLPCs, and we defined a NT-rich microenvironment for LLPC survival in nasal polyps. LLPCs are resistant to glucocorticoids, irradiation, conventional immunosuppressive, and cytotoxic drugs, and B cell depletion therapies.637 It is possible that LLPCs in nasal polyps may contribute to glucocorticoid treatment resistance. Novel therapies targeting LLPCs and their specific survival niches may improve the treatment outcome of nasal polyps.
 XML Download
XML Download