

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Post-transcriptional regulations refer to the processes that RNA transcripts are subjected to between transcription and translation, namely 5′-capping, splicing, polyadenylation, RNA modification, etc. These processes greatly impact the expression of genes (Fig. 1A). Particularly, recent studies have convincingly demonstrated that post-transcriptional regulations play critical roles in controlling cellular biology and are associated with various diseases.1-4 However, our knowledge on the mechanistic integration of the control of post-transcriptional regulations to cellular signaling pathways is still largely lacking.
Fig. 1
Overview of post-transcriptional regulations in eukaryotic cells. (A) Co-transcriptional events and transcriptional termination. Post-transcriptional processing, i.e. splicing, polyadenylation, AS, and APA occur co-transcriptionally. These post-transcriptional events can produce transcript isoforms from genes and contribute to the diversity and dynamics of the transcriptome and the resulting proteome. (B) The 5 different types of AS events. (C) The 2 types of APA events. The 3′-UTRs serve as binding platforms of various regulatory RBPs and miRNAs (upper). UTR-APA, since most of the alternative PASs are proximal, 3′-UTRs are often shortened, resulting in the production of transcripts that can escape the regulation of those regulatory factors. The 2 types of CR-APA (lower).
P, promoter; Pol II, polymerase II; PAS, poly-A signal; 5′-P, 5′ phosphate group; Xrn2, 5′-3′ exoribonuclease 2; UTR, untranslated region; CDS, coding DNA sequences; CR, coding region; APA, alternative polyadenylation; SS, splice site; RBP, RNA-binding protein.
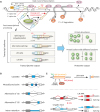
Mammalian target of rapamycin (mTOR) is a serine/threonine kinase that forms two functional complexes known as mammalian target of rapamycin complex 1 (mTORC1) and mammalian target of rapamycin complex 2 (mTORC2). Raptor and Rictor are the specific components for mTORC1 and mTORC2, respectively. Although mTORC2 has been functionally associated with cell survival, metabolic control and cytoskeleton organization, its regulations and functions still await more thorough studies to be revealed.5 On the other hand, mTORC1 has been extensively studied in the past few decades due to its important roles in regulating various anabolic and catabolic processes, as well as its involvement in the pathogenesis of cardiovascular diseases, diabetes, and many types of cancers.5-7 As a master regulator of various metabolic processes, mTORC1 is sensitive to growth factor signaling, nutrient and oxygen availability, and intracellular energy status. And many studies have shown that mTORC1 regulates various cellular metabolism pathways through modulating the translation and/or transcription activities of key downstream effectors.5,7-9 Intriguingly, recent studies have also revealed mTORC1's involvement in gene expression regulation at the post-transcriptional level.
In this review, we use mTORC1 signaling pathway as an example to highlight the significant roles of post-transcriptional regulations in cellular gene expression regulation. Below, we first describe recent understandings of post-transcriptional regulations in the field, focusing on splicing and polyadenylation, which are more extensively studied than other processes. We also briefly discuss the centrality of mTORC1 in the regulation of various biological processes through translational and transcriptional controls. Then, we present recent discoveries of mTORC1's roles in post-transcriptional regulations and their physiological outcomes. Finally, we share our opinion regarding the implications of these recent discoveries on our approach towards scientific studies involving transcriptome datasets.
POST-TRANSCRIPTIONAL REGULATIONS AND THEIR ROLES IN GENE EXPRESSION REGULATION
Splicing and alternative splicing (AS)
Splicing is a critical mRNA maturation process in eukaryotic cells in which intervening sequences (introns) are cut out from the nascent transcript and exon sequences are pieced together by the spliceosome to form uninterrupted coding DNA sequences (CDS) and untranslated region (UTR) sequences for proper protein translation. This process occurs co-transcriptionally and is dependent upon the actions and activities of RNA polymerase II (Pol II) (Fig. 1A).10-12 In AS, certain exons, or part of certain exons, are alternatively included in the mature transcripts in different biological contexts. AS occurs in about 95% of multi-exon genes in human transcriptome.13 The regulation of AS is dependent upon the coordination of relevant trans-acting RNA binding proteins (RBPs) and the cis-acting elements surrounding the alternative exons. A series of RBPs such as serine arginine (SR) proteins and heterogeneous nuclear ribonucleoproteins (hnRNPs) that can affect the molecular actions of the spliceosomal complexes play important roles in the regulation of general RNA splicing and AS.14,15 There are generally 5 basic types of AS16,17: cassette type (or exon inclusion/skipping type) occurs when the 3′-splice site of the downstream intron is used for splicing, skipping over an entire exon; mutually exclusive type occurs when 2 cassette exons are mutually exclusively selected for splicing; alternative 5′-splice site type occurs when a different 5′-splice site is defined and used for splicing; alternative 3′-splice site type occurs when a different 3′-splice site is recognized; finally, intron retention occurs when neither the 5′-splice site nor the 3′-splice site of an intron is used for splicing, leaving that particular intron in the final mRNA transcript (Fig. 1B). Since AS alters the sequence of the resulting mature mRNA, it can impact the expression and function of the gene. For example, given that 5′-UTRs often contain sequence elements or secondary structures that can affect ribosome loading, AS of exons in the 5′-UTRs may alter the translation efficiency of the transcripts.18,19 Moreover, AS of exons in the CDS may alter the function of certain domains or affect the folding of proteins, leading to changes in the characteristics of the protein products translated from the transcripts. These AS events increase the capacity of functional proteome.17 And more importantly, AS events can have significant physiological impacts without necessarily affecting the transcript expression level of genes. Indeed, multiple biological contexts and diseases, e.g. cancers, have been associated with transcriptome-wide AS events.1,2 These demonstrate the importance of studying the roles of AS events in cellular gene expression regulations.
Transcriptional termination, polyadenylation, and alternative polyadenylation (APA)
The termination of the transcription activity by RNA Pol II involves a sequence of molecular events. Towards the end of transcription, the nascent transcript undergoes endonucleolytic cleavage to be released and polyadenylated while Pol II continues with transcription. Then, Pol II is released from the DNA for recycling and allow for the next rounds of transcription (Fig. 1A). For cleavage and polyadenylation, namely the maturation of the 3′ end of mRNA molecules (sometimes simply referred to as polyadenylation), Pol II first reaches and transcribes sequence elements that recruit the formation of the 3′-end processing complex, which include a poly-A signal (PAS, most commonly AAUAAA, AUUAAA, and several other variants), and often a U-rich auxiliary upstream element (USE) and an U-rich or AU-rich downstream element (DSE). These signal the recruitment of trans-acting factors such as the PAS-binding cleavage and polyadenylation specificity factors (CPSFs) and the DSE-binding cleavage stimulation factors (CSTFs) to bind to the nascent transcript to catalyze an endonucleolytic reaction at a CA dinucleotide that is usually 15–30 nucleotides downstream of the PAS. Then, poly-A polymerase adds a stretch of untemplated adenosines, the poly-A tail, to the 3′-end of the transcript from the cleavage site. The poly-A tail is needed for downstream metabolism of the mature mRNA transcript including nuclear export of mRNA, translation, localization, and stability.20-22 As for the release and the termination of the transcription reaction of Pol II, 5′ to 3′ exonucleases are recruited to attack the 5′-end generated by the cleavage during polyadenylation process, which is unprotected by a 5′-cap. The exonucleases (e.g. Xrn2, 5′-3′ exoribonuclease 2) then chase down Pol II along their substrate and finally displace Pol II from the transcription bubble to terminate the transcription reaction.23-25
Thus, termination of transcription reactions is dependent upon the occurrence and “strength” of the cis-acting elements that signal for polyadenylation (the USE-PAS-DSE pattern). Interestingly, at least 70% of human genes are predicted to possess two or more such cis-acting elements.26 And, when more than one PASs in a gene are capable to be utilized for 3′-end processing, APA occurs. The regulation of APA, like AS, is determined by the coordination of various trans-acting factors and cis-acting elements surrounding the PAS and alternative PAS.3,4 The expression levels and activities of a number of RBPs have been shown to be able to affect APA at a transcriptome-wide level due to their roles in interacting with 3′-end processing factors and/or the cis-acting elements near PASs; these factors include the components of the 3′-end processing complex such as CPSFs and CSTFs, as well as other polyadenylation-associated RBPs such as cytoplasmic polyadenylation element binding protein 1 (CPEB1) and poly(A) binding protein nuclear 1 (PABPN1).27-32 Generally, there are 2 types of APA: UTR-APA and coding region (CR)-APA.33 3′-UTR of genes is often longer than CDS and contains binding sites for microRNAs and regulatory RBPs. It also contains alternative PASs. Most of the alternative PASs in 3′-UTRs are upstream of the canonical or annotated PASs (and in this scenario, the alternative PASs are termed proximal PASs and the canonical PASs are termed distal PASs, based on their relative distance to the stop codon). Thus, when these alternative PASs are utilized for polyadenylation, namely when UTR-APA occurs, in most cases, 3′-UTRs are shortened. This phenomenon is referred to as 3′-UTR shortening. As UTR-APA alters the availability of these regulatory cis-acting elements on the mature transcripts, it can affect the behaviors of mRNA transcripts, including translation efficiency, localization, and stability, without changing the coding capacity of the transcripts (Fig. 1C).3,4,26,34,35 For example, PAX3, one of the master transcription regulators of the myogenic transcriptional network, has a miR-206 binding site in its 3′-UTR. It has been observed that among different muscle cell types, the ratios of PAX3 UTR-APA isoforms (thus the ratio of transcripts being able to be regulated by miR-206) differ. This results in varying degrees of protein translation efficiency for PAX3 transcripts in the different muscle cell types, which can help explain the varying differentiation patterns in these distinct muscle cell types.36 It has also been shown that in neurons, where accurate localization of gene expression is crucial for proper cellular functions, the isoforms of hundreds of genes are differentially localized based on the UTR-APA events in their 3′-UTRs.37 These highlight the crucial role of UTR-APA events in determining the fate of mRNA transcripts. Furthermore, the 3′-UTR of an mRNA transcript has also been shown to serve as a molecular scaffold for protein-protein interactions, particularly, immediate interactions between the nascent protein synthesized from the mRNA transcript and its binding partners. For example, the 3′-UTR of the membrane protein CD47, can recruit protein complexes including ELAV like RNA binding protein 1 (ELAVL1, or HuR) and SET nuclear proto-oncogene (SET), allowing immediate interaction of these proteins with the nascent CD47 protein. This molecular event leads to the efficient translocation of CD47 to the plasma membrane. Upon UTR-APA, CD47 transcript can no longer recruit the binding partners for plasma membrane localization. The CD47 protein produced from CD47 transcript with UTR-APA localizes to endothelium reticulum, instead.38,39 As for CR-APA, it occurs when alternative PASs in the upstream intronic regions are utilized for polyadenylation. It is thus also sometimes referred to as intronic APA. Once a transcript is truncated, the transcript would lose the coding capacity of a chunk of polypeptide on the C-terminal end. The resulting protein product may thus function differently. Moreover, the truncated transcript would be differentially regulated compared to the full-length counterpart as it would possess a completely different 3′-UTR that originates from the intron region downstream of the alternative PAS. There are 2 types of CR-APA; the mechanism of both types of CR-APA involves the interplay between splicing and polyadenylation.3,33,40 The first type of CR-APA occurs when the splicing of a PAS-containing intron is inhibited, and the 3′-end processing complex outcompetes the splicing machinery, leading to truncation of the transcript at that intron. The second type of CR-APA occurs when a cryptic exon that is followed by a PAS is utilized for splicing. Due to the presence of the PAS, this “alternative splicing” event leads to the truncation of the mRNA transcript (Fig. 1C). While the studies in the field focusing on CR-APA is relatively few, one of the first and most famous identified APA events belongs to this category. In resting B lymphocytes, immunoglobulin heavy constant mu (IGHM) gene is transcribed to full-length, which includes the last 2 exons that encode a transmembrane domain, leading to the expression of membrane-bound antibody chain. And upon B lymphocyte activation, CR-APA of IGHM transcript occurs, causing the two exons encoding the transmembrane domain to be excluded from the final mRNA transcript. This leads to the production of secreted form of immunoglobulin M antibody chain.41 Another example of CR-APA is the mitogen-activated protein kinase (MAPK) interacting serine/threonine kinase 1 (MKNK1) gene. The CR-APA event of MKNK1 gene results in the loss of 89 amino acids at the C-terminal end of the kinase and the addition of 12 amino acids encoded by the intronic sequence. It has been shown that this truncated version of MKNK1 has a higher kinase activity compared to its full-length counterpart.42 Moreover, intriguingly, a recent study found that in chronic lymphocytic leukemia cells, a number of tumor suppressor genes undergo CR-APA and consequently lose their tumor-suppressing activities. Some of the truncated proteins even seemed to act in an oncogenic manner.43 Together, all these previous findings show that APA can drastically affect gene expression and steer cellular biology independent of the transcription regulation of genes.
mTORC1 REGULATES VARIOUS BIOLOGICAL PROCESSES THROUGH TRANSLATIONAL AND TRANSCRIPTIONAL CONTROLS
It is generally understood that the activated mTORC1 promotes key anabolic processes including protein synthesis, lipid synthesis, nucleotide synthesis, biogenesis of organelles, and that it inhibits certain catabolic processes such as autophagy. As a result, activation of mTORC1 leads to cellular growth and proliferation.44 On the other hand, decreased mTORC1 activity, e.g. when cells experience nutrient deprivation, can activate autophagy and lower mitochondrial membrane potential in cells so that cellular energy may be conserved.45,46 Moreover, activation of mTORC1 has also been shown to promote angiogenesis and inflammation in certain tissues.47-49 It is thus not surprising that mTORC1 inhibition has been proposed and used as a treatment for several types of cancers and atherosclerosis.50,51
mTORC1 is negatively regulated by tuberous sclerosis complexes (TSC1/2), which are downstream of and can be regulated by several major cellular signaling pathways including phosphoinositide-3-kinase/protein kinase B signaling pathway, AMP-activated protein kinase signaling pathway, MAPK/extracellular-signal-regulated kinase pathway, etc.5
Many studies have shown that mTORC1 regulates its various downstream pathways through translational and transcriptional controls. One of the most well-known functions of mTORC1 is the direct phosphorylation of ribosomal protein S6 kinase beta-1 (S6K1) and eukaryotic initiation factor 4E-binding protein 1 (4E-BP1) (Fig. 2). These lead to the increase of protein synthesis efficiency, particularly for transcripts containing 5′-terminal oligopyrimidine tract or 5′-pyrimidine-rich translational element in their 5′-UTRs.9,52,53 Moreover, mTORC1 phosphorylates protein phosphatase 2A (PP2A) and transcription initiation factor 1A (TIF-1A) to increase the transcription of ribosomal RNAs, enabling efficient cellular protein synthesis (Fig. 2).54 One of the genes whose protein production is elevated by activated mTORC1 is the activating transcription factor 4 (ATF4). Through the transcription regulation of ATF4 on several genes involved in de novo purine synthesis, mTORC1 can promote purine synthesis in cells.55 mTORC1 also positively regulate lipid and sterol biosynthesis by activating transcription factors such as sterol regulatory element binding protein 1 (SREBP1), peroxisome proliferator-activated receptor γ (PPARγ), and PPARγ coactivator 1 alpha (PGC-1α), which control the transcription of genes involved in de novo lipid synthesis as well as lipid and cholesterol homeostasis.56 Through PGC-1α, mTORC1 also controls mitochondrial biogenesis and oxidative metabolism (Fig. 2).57 On top of these, it has been shown that mTORC1 can promote angiogenesis by increasing the translation activity of hypoxia-inducible factor 1 subunit alpha (HIF-1α) gene, a transcription factor that regulates the expression of several angiogenic growth factors including vascular endothelial growth factor.47 Furthermore, through regulating the activities of inflammatory transcription factors such as nuclear factor kappa B (NF-κB) and signal transducer and activator of transcription-3 (STAT3), mTORC1 can also control pro- and anti-inflammatory responses in blood cells, depending on cellular contexts.49 Again, as these examples demonstrate, it has been well-established that mTORC1 regulates a variety of cellular processes through controlling the translation and transcription activities of its downstream effector genes (Fig. 2).
Fig. 2
Illustration of mTORC1's translational and transcriptional controls over various metabolic pathways and physiological outcomes. The activation of mTORC1 not only leads to the upregulation of cellular translation activity, but also regulates various metabolic pathways through controlling transcription networks.
mTORC1, mammalian target of rapamycin complex 1; HIF-1, hypoxia-inducible factor 1; Nrf2, nuclear factor erythroid 2-related factor 2; HSF1, heat shock factor 1; NF-κB, nuclear factor kappa B; STAT3, signal transducer and activator of transcription-3; ATF4, activating transcription factor 4; PPAR, peroxisome proliferator-activated receptor γ; SREBP-1, sterol regulatory element binding protein 1; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1 alpha; mTOR, mammalian target of rapamycin; PRAS40, proline-rich Akt substrate of 40 kDa; PP2A, protein phosphatase 2A; TIF-1A, transcription initiation factor 1A; Atg, autophagy-related; ULK, Unc-51-like kinase; 4E-BP, 4E-binding protein; eIF4G, eukaryotic translation initiation factor 4G.
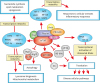
mTORC1-COORDINATED POST-TRANSCRIPTIONAL REGULATIONS AND THEIR PHYSIOLOGICAL OUTCOMES
mTORC1-mediated polyadenylation events and profiling technologies of polyadenylation
With the advent of high-resolution deep sequencing technologies and numerous bioinformatics pipelines for data analyses, transcriptome-wide gene expression can be understood and alternative processing of gene products can be profiled. Transcriptome-wide studies on AS and APA have thus been conducted routinely in numerous biological systems and in various physiological contexts. A number of studies reporting mTORC1's role in transcriptome-wide post-transcriptional regulations have also emerged.
In mouse embryonic fibroblast cell (MEF) model, a genetic activation of mTOR by knocking out Tsc1 (Tsc1−/−) showed an increase of transcriptome-wide UTR-APA events. Interestingly, the majority of the transcripts with shortened 3′-UTR by UTR-APA did not show significant overall transcript level changes. Importantly, polysome profiling for actively translating transcripts indicated that 3′-UTR shortening by APA in a transcript promotes the efficiency of protein synthesis. Thus, this transcriptome-wide 3′-UTR shortening phenomenon in the mTOR-activated transcriptome highlights the fact that traditional RNA-Seq analyses that only focus on gene expression level profiling can miss a significant portion of expression-regulating events in the transcriptome by mTORC1 (Fig. 3).58
Fig. 3
Illustration of how mTORC1-mediated post-transcriptional regulations play a role in controlling various cellular pathways and physiological outcomes. Recent studies have shown that mTORC1 controls the AS and APA of select genes, affecting their expressions. These can lead to changes in cellular biology, e.g. proliferation.
mTORC1, mammalian target of rapamycin complex 1; AS, alternative splicing; APA, alternative polyadenylation; mTOR, mammalian target of rapamycin; PRAS40, proline-rich Akt substrate of 40 kDa; U2AF1, U2 small nuclear RNA auxiliary factor 1; S6K1, S6 kinase beta-1; SRPK2, serine and arginine rich splicing factor protein kinase 2; SR, serine arginine; UTR, untranslated region; DGE, differential gene expression; ER, endoplasmic reticulum.
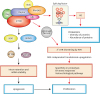
In general, most of the bioinformatics tools for UTR-APA profiling, including the one used in the above study, take advantage of the highly quantitative nature of RNA-Seq for the measurement of UTR-APA events by comparing the ratios of the read density of the long form-only regions and the read density of the regions common to both long and short transcripts between 2 samples. Changes of the ratios between 2 samples indicate the occurrence of an UTR-APA event. A major issue with this RNA-Seq-based approach for UTR-APA analysis is that the measurement relies on the accurate information on the locations of the 3′-ends of the short transcripts. UTR-APA events involving unannotated short transcripts can be missed, while UTR-APA events involving annotated yet biologically irrelevant (not expressed in the cell line or tissue under study) short transcript isoforms can be falsely identified.33,59,60
An attempt to overcome this existing issue on UTR-APA analysis was made by integrating two sequencing technologies together. In that follow-up report, poly-A site sequencing (PAS-Seq) was performed on 2 MEF models with basal (WT) and hyper mTOR (Tsc1−/−) activity. PAS-Seq is a sequencing technique that focuses the sequencing depth of RNA-Seq onto the polyadenylated sites of mRNA transcripts for a more confident and experimentally-proven profiling of PASs used for polyadenylation in the biological system. By integrating the PAS-Seq data sets and the previous RNA-Seq results, the confidence and accuracy of identification and quantification of biologically relevant UTR-APA events in the mTORC1-activated transcriptome were both elevated. Although this new method requires that the same samples must be sequenced by both RNA-Seq and PAS-Seq, which can be costly and prevents this method to be applied to old RNA-Seq data that lack the corresponding PAS-Seq information, this method did allow the identification of new biological pathways regulated by mTORC1-mediated UTR-APA events.61 Therefore, with ever-improving sequencing technologies and bioinformatics tools, the role and impact of mTORC1 in transcriptome-wide UTR-APA regulations have become clearer.
Physiological outcomes of mTORC1-mediated APA
Since 3′-UTR of genes contain many microRNA and RBP-binding sites, 3′-UTR shortening would allow transcripts to escape the regulation of these trans-acting factors. In a similar context, siRNAs targeting 3′-UTR have been shown to be ineffective upon mTOR-driven shortening of select transcripts, suggesting that APA modulates diverse regulatory activities that may happen through 3′-UTR.58 As such, 3′-UTR shortening has been shown to alter the stability, localization, and translation efficiency of mRNA transcripts.3,4,26 In select cases of the mTORC1-activated 3′-UTR shortening events, a significant increase in protein synthesis without significant changes in their mRNA transcript levels was observed. Pathway enrichment analysis on these 3′-UTR-shortened genes revealed that ubiquitin-mediated proteolysis pathway is the most targeted pathway by mTORC1-mediated 3′-UTR shortening (Fig. 3).58 Thus, with UTR-APA analysis on mTORC1-activated transcriptome, a pathway activated by mTORC1 that was previously not associated with mTORC1 activity was revealed. Moreover, with the integration of PAS-Seq in our UTR-APA analysis, it was further revealed that mTORC1 regulates the 3′-UTR shortening of several transcription factors, suggesting that mTORC1 may have a role in controlling various transcription networks through its regulation of UTR-APA. For example, it was demonstrated that mTORC1 upregulates CCAAT/enhancer binding protein gamma (CEBPG) through 3′-UTR shortening; this upregulation of CEBPG by mTORC1 is critical in protecting cells against endothelial reticulum stress (Fig. 3).61
mTORC1-mediated AS
Passacantilli et al.62 reported that upon chemical mTOR inhibition, extensive changes in the transcriptome were observed in Ewing sarcoma cells using microarray analysis. Particularly, 1,440 AS events were detected in 918 genes.62 These data were obtained by microarray technology, which can only identify previously annotated AS events. This suggests that we could expect to observe more mTOR-mediated AS events if unbiased sequencing technologies such as RNA-Seq are used for analyzing these biological samples. Nonetheless, their work demonstrates that mTOR activity indeed regulate transcriptome-wide AS events. Furthermore, Lee et al.63 also reported that chemical inhibition of mTORC1 leads to malfunction in the splicing of select genes. With improper splicing, introns are retained in these genes, activating nonsense-mediated pathway for the degradation of the transcripts (Fig. 3).63
Physiological outcomes of mTORC1-mediated AS
Lee et al.63 demonstrated that, through S6K1 phosphorylation, mTORC1 regulates the activity of serine and arginine rich splicing factor protein kinase 2 (SRPK2), a key regulator of a series of splicing factors. When mTORC1 is downregulated, SRPK2 activity is inhibited, preventing the proper function of downstream splicing factors, causing splicing dysregulation and thus downregulation of a number of lipogenic genes (Fig. 3).63 This study showed that mTORC1, through regulating the activities of splicing regulators, can modulate gene expression at the level of splicing regulation. Passacantilli et al.62 also demonstrated that the transcriptome-wide AS events they observed in embryonic stem cells upon mTOR inhibition contribute to drug resistance in that particular cancer cell line.62 Moreover, recently, it was shown that cellular mTOR activity regulates the expression of U2AF1 isoforms (U2AF1a v. U2AF1b), a critical splicing factor determining 3′-splice site. One striking molecular outcome of this differential U2AF1 isoform expression is the changes of AS in 5′-UTR of many genes that significantly affect translation efficiency.64 Thus, these reports not only reveal mTORC1's role in splicing regulation, but that these mTORC1-mediated splicing regulations have significant physiological impacts.
Summary
Taken together, these studies add post-transcriptional regulation as another layer of gene expression regulation to our understanding of the multi-faceted functions of mTORC1 (Fig. 3). Moreover, analyses of mTORC1-mediated post-transcriptional regulations in various cellular contexts have enabled researchers to not only gain new mechanistic insights into mTORC1's control over previously associated biological processes, but also to discover new cellular signaling pathways that are regulated by mTORC1 and make mechanistic connections that were previously masked due to a lack of analyses focusing on post-transcriptional regulations.
CONCLUDING REMARKS
It has been a common knowledge that if we apply the simplistic understanding of the central dogma and the one-gene one-enzyme theory in molecular biology to our understanding of gene expression regulation in cells, we may fail to fully capture the complex landscape of gene expression regulations and the dynamics of functional proteome in cells. As discussed above, the regulations on mRNA transcripts that occur post-transcriptionally, namely post-transcriptional regulations, play important roles in the diversity and dynamics of the functional proteomes of biological systems. The physiological impacts of these post-transcriptional regulations can be quite significant and should not be neglected when we study the expression of genes in cells. Nonetheless, most commonly, when researchers perform transcriptome profiling with technologies such as RNA-Seq, only transcript level analyses are carried out. Moreover, most, if not all, transcript-based biomarkers presently available are designed according to the transcript level profiling of the samples. Indeed, mRNA transcript level changes can suggest physiological outcomes due to protein level changes, yet often times transcript level changes do not lead to corresponding protein level changes. And intriguingly, for many of such cases, post-transcriptional regulations are the reason for these seeming discrepancies.65 Therefore, given that many genes have been shown to display drastically different functions through AS or APA without changing their transcript levels necessarily, and that multiple user friendly and free bioinformatics tools are currently available online,33 analyses on post-transcriptional regulations with transcriptome profiling datasets should be performed routinely in order to capture the fuller picture of gene expression regulations in the biological systems of interest.
In this review, we have discussed the significance of AS and APA events in cellular biology in the context of mTORC1 signaling. We show that the regulation of post-transcriptional events by mTORC1 is just as extensive and important to cellular biology as its regulations of transcriptional and translational events. This not only helps cement the role of post-transcriptional regulation as an important layer of gene expression regulation, it also establishes a mechanistic link between a well-studied and high-profile cellular signaling pathway, mTORC1, to the regulation of post-transcriptional events, as well as the physiological outcomes of these events. On the other hand, we believe that post-transcriptional regulations can also play as important and extensive of a role in other cellular signaling contexts as observed in mTORC1 signaling pathway. With the rising awareness of how post-transcriptional regulations can dictate and steer cellular biology, and the increasing availability and sophistication of technologies to study post-transcriptional events, we expect to see more and more reports on the key roles of post-transcriptional events in various cellular signaling pathways and the pathogenesis of diseases in the near future.
Furthermore, apart from the two types of post-transcriptional events highlighted in this review (AS and APA), there are other types of regulations at the mRNA transcript level that can also be studied with transcriptome-profiling data. They all add to the richness of information one can obtain on top of transcript level analysis with transcriptome data, given that suitable bioinformatics tools are available. One example would be alternative transcription start site events, where the same gene uses different transcription start sites for transcription under different circumstances.66 The consequences of alternative transcription start site events include alternative 5′-UTR composition, potentially resulting in changes in the fates of the final mRNA transcripts, as well as changes in the N-terminal ends of proteins, potentially leading to the inclusion or exclusion of signal peptides or ubiquitin sites in the final protein products. Another common type of regulation on mRNA transcripts is RNA modification/editing, where the insertion, deletion, and/or substitution of nucleotides is carried out on a transcribed RNA molecule by a set of enzymes. These editing events can lead to changes in the coding of amino acids of the final protein product, or changes in the binding sites of trans-acting factors, alternating the regulations on the mRNA transcripts by these factors.67 Indeed, these regulations on mRNA molecules can have exciting biological consequences. However, they are not very well explored in the field yet, at least not systematically. Thus, with single-nucleotide resolution sequencing experiments being routine for examining the transcriptomes of biological samples nowadays, researchers can and should perform analyses on post-transcriptional regulations and events on top of transcript level analysis to be better informed when studying the biological phenomena of interest.
 XML Download
XML Download