

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION TO CARDIOVASCULAR DISEASE-ASSOCIATED MACROPHAGE INFLAMMATION
Cardiovascular disease (CVD) remains the leading cause of death globally and in the United States.1 CVD encompasses a wide range of diseases that affect the heart and blood vessels, including but not limited to conditions such as coronary heart disease, heart failure, and hypertensive heart disease. While the scope of CVD is broad, there is a gradient in terms of the burden of various types of CVD, with atherosclerotic vascular disease currently dominating worldwide.2 Atherosclerosis is marked by immune activation and lipid accumulation in the arterial wall. It is also associated with risk factors such as high blood pressure, smoking, obesity, and high cholesterol.3 The pathogenesis of atherosclerosis involves a complex interplay of multiple cell types.4 In particular, macrophages are central protagonists in the genesis and progression of atherosclerotic CVD. During atherogenesis, infiltration of macrophage precursor monocytes into atherosclerotic lesions and in response to the retention of sub-endothelial lipoprotein are important components of the progression from pathologic intimal thickening to late fibroatheroma.5 Macrophages also play a critical role in cardiac repair and recovery following atherothrombotic myocardial infarction.6 In particular, heterogeneous macrophage populations carry out distinct functions during cardiac repair following cardiac insult.78 Numerous regulatory mechanisms govern macrophages and their inflammatory functions. Non-coding RNAs (ncRNAs), historically considered unnecessary products of so-called “junk” DNA, have emerged as key contributors to macrophage regulation.
ncRNAs AND lncRNAs
Modern genomic and transcriptomic approaches have opened our eyes to the potential role of ncRNAs in the nuanced regulation of gene expression, cell function,9 and disease pathogenesis.10 Initially characterized in bacteria,11 the field quickly expanded to identify roles played by ncRNAs in mitochondrial12 and macrophage13 regulation. ncRNAs are categorized based on size. Small ncRNAs are less than 200 nucleotides in length, while long non-coding RNAs (lncRNAs) are greater than 200 nucleotides. The size of ncRNAs also generally correlates with their function. For example, microRNAs (miRNAs) are 20–24 nucleotides in length and bind to the 3' untranslated region of messenger RNA (mRNA), thereby sterically blocking translation and increasing mRNA degradation.14 On the longer side of the spectrum, lncRNAs are able to interact with DNA, RNA, and proteins to exert cellular regulation at multiple levels, including chromatin remodeling,15 mRNA splicing,16 mRNA translation,17 and multi-protein complex assembly.18 The enhancer RNA subset of lncRNAs are less than 2,000 nucleotide-long transcripts that are produced from enhancer domains to regulate the transcription of target genes19 or nearby genes through diverse mechanisms.20 Importantly, many lncRNAs fall within intergenic regions of the genome (lincRNAs), which are under less selective pressure than genic sequences. These lincRNAs may thus have opportunities for functional diversification compared to their genic counterparts, and could encode species-specific functions.21 In humans, ncRNAs are implicated in cardiogenesis,22 ventricular remodeling after myocardial infarction,23 and wound healing.24
In this brief review, we discuss the role of select ncRNAs, including their potential role in myeloid cell metabolism (immunometabolism). Given the relatively early state of the field, we chose to focus on specific ncRNAs that either have been extensively validated by independent laboratories or have significant potential for future study. We focused on the aspects of these ncRNAs that affect macrophage inflammatory function in the context of atherosclerosis and myocardial infarction. It is also important to note that ncRNAs are expressed in non-myeloid cells; however, the focus of this review is myeloid cells and macrophages. For example, the complexity and heterogeneity of macrophages require dynamic metabolic reprogramming.25 In recent years, novel experimental approaches have enabled researchers to identify specific metabolic variations and their direct connections to immune cell activation. In lipopolysaccharide-activated macrophages, dendritic cells, and activated B and T cells, there is often evidence of enhanced glycolysis following activation.25 In atherosclerosis and related cardiovascular diseases, the integration of excess lipid metabolism and myeloid metabolic processes provides a unique set of circumstances that are likely under the control of lncRNAs.
SPECIFIC ncRNAs OF INTEREST FOR MACROPHAGES AND CARDIOVASCULAR DISEASE
Below we highlight specific ncRNAs of interest, as also shown in Fig. 1. The lncRNA ANRIL (CDKN2BAS) is remarkable in its capacity to regulate genes that have been extensively linked to glucose and fatty acid metabolism.26 Like protein-coding genes, ANRIL is alternatively spliced into multiple linear and circular forms in a tissue-specific manner. In macrophages, ANRIL expression triggers increased glycolysis and induces apoptosis.27 ANRIL is induced by glucose uptake28 and proinflammatory factors such as interferon gamma29 and tumor necrosis factor alpha (TNF-α).30 ANRIL may work through numerous mechanisms, including as an epigenetic modifier or miRNA sponge.31 ANRIL has the capacity to regulate interleukin (IL)-6 cytokine production, for example through the transcription factor Yin Yang 1 (YY1), a component of the nuclear factor-κB pathway.30 Moreover, ANRIL is associated with hematopoietic stem cell proliferation, a key component of the myeloid response after myocardial infarction.32 To maintain homeostasis, excess hematopoietic stem cell proliferation is partially controlled by the tumor suppressor genes cyclin-dependent kinase 2A (CDKN2A) and 2B (CDKN2B).33 Located within a single topological domain, ANRIL negatively regulates CDKN2A/B expression via chromatin remodeling.26 In THP1 macrophage-derived foam cell lines, ANRIL suppressed atherosclerotic-like inflammatory responses and promoted cholesterol efflux.34 ANRIL was shown to regulate ADAM10, a disintegrin and metalloproteinase domain-containing protein, that has been shown to be involved in neuroinflammation35 and expressed at high levels in atherosclerosis.36 The overexpression of ANRIL was found to result in methylation of the ADAM10 gene, which has been shown to inhibit atherosclerotic inflammation.34 Importantly, single-nucleotide polymorphisms within the ANRIL sequence are correlated with an increased risk of coronary artery disease and type 2 diabetes.3738 It will be interesting to determine whether ANRIL expression alters myeloid function through metabolic manipulation after ischemic insults.
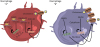 | Fig. 1Select ncRNAs of potential significance in the regulation of macrophage function during cardiovascular disease. ncRNAs enact multiple regulatory functions in macrophages. Depicted in the schematic are the potential roles of the ncRNAs NEAT1, GAS5, DAPK-IT1, miR-33, ANRIL, and MeXis. NEAT1 and GAS5 acts as sponges of miR-342-3p and miR-135a, respectively, leading to the upregulation of inflammatory cytokines. DAPK-IT1 and miR-33 both inhibit expression of the ABCA1 gene, leading to reduced expression of ABCA1 cholesterol transporters. ANRIL and MeXis increase macrophage cholesterol efflux, with Mexis working specifically at the ABCA1 gene locus, resulting in increased ABCA1 expression.ncRNA, non-coding RNA.
|
Another important lncRNA involved in cholesterol metabolism in macrophages is MeXis, which lies in close proximity to the locus of the cholesterol-efflux gene ABCA1.3940 MeXis enhances binding of the transcriptional coactivator DDX17 to enhancer regions in the ABCA1 gene locus, thereby increasing nuclear receptor LXR-mediated gene expression of ABCA1 in macrophages. Single-nucleotide polymorphism variants within the MeXis locus are correlated with an increased risk of coronary artery disease in humans.39 Importantly, MeXis is preferentially expressed in macrophages in the heart and kidney, whereas a distinct lncRNA, LeXis, is preferentially expressed in the liver.39 MeXis and LeXis are interesting in that they reveal a mechanism by which cells respond uniquely to the same stimulus, resulting in tissue-specific changes in metabolic regulation. Consequently, MeXis expression leads to increased levels of intracellular cholesterol in cardiac macrophages, in turn inducing inflammatory pathways that are implicated in myocardial infarction.
The lncRNA NEAT1 is involved in cholesterol metabolism and atherosclerosis development. Since its discovery, NEAT1 expression has been discovered in a number of diseases, including lupus41 and several disparate cancers.4243 A lncRNA screen of peripheral blood mononuclear cells revealed differential expression of NEAT1 in patients following myocardial infarction.44 NEAT1 was the most highly expressed and subsequently deregulated lncRNA identified. Multivariate statistical analysis revealed that NEAT1 levels were correlated with post-myocardial infarction status, independent of statin intake, left ventricular ejection fraction, low-density lipoprotein (LDL) or high-density lipoprotein cholesterol, age, diabetes, and smoking.44 NEAT1 has also been shown to be upregulated in THP1 macrophages stimulated by oxidized LDLs (ox-LDLs).45 Knockdown of NEAT1 in these THP1 cells repressed the formation of foam cells triggered by ox-LDLs.45 NEAT1 functions partly by acting as a sponge for miR-342-3p, as depicted in Fig. 1. Repression of NEAT1 and overexpression of miR-342-3p inhibited lipid uptake in THP1 cells. Therefore, it appears that NEAT1 promotes inflammation and lipid uptake through miR-342-3p repression.45 NEAT1 has been experimentally shown to inhibit another miRNA, miR-128. Analysis of RAW264.7 cells stimulated by ox-LDL showed increased NEAT1 expression in combination with decreased miR-128 expression.46 Knockdown of NEAT1 in the same cells repressed foam cell formation and downregulated IL-6, IL-1β, and TNF-α.46 As with miR-342-3p, overexpression of miR-128 inhibited atherosclerotic development triggered by NEAT1.46 In mice, Neat1 enhanced activation of the NLRP3, NLRC4, and AIM2 inflammasomes by stabilizing inflammasome caspase tetramers.47 With all of these considerations in mind, further research into NEAT1 could lead to the identification of possible therapeutic targets in atherosclerotic development.
In recent years, miR-33 has emerged as a significant regulator of macrophage function and cholesterol efflux.48 In humans, miR-33 is co-expressed with the SREBF1/2 genes, which code for transcription factors that regulate fatty acid synthesis and uptake.4950 miR-33 expression inhibits the expression of ABCA1 and ABCG1, two key transporters involved in cholesterol efflux in macrophages, while SREBF1/2 regulates cholesterol biosynthesis and cellular uptake.51 Therefore, miR-33 and SREBF1/2 co-expression is dependent on intracellular cholesterol concentrations, with low-sterol conditions leading to the upregulation of cholesterol biosynthesis through SREBF1/2, and reduced cholesterol efflux through miR-33. Moreover, miR-33 can regulate mitochondrial respiration and metabolism by inhibiting expression of the PGC-1α, PDK4, and SLC25A25 proteins.49 As oxidative phosphorylation is a downstream function of PGC-1α expression, anti-miR-33 treatment was found to lead to increased mitochondrial biogenesis and efficient ATP production. miR-33 has also been shown to regulate autophagy and lipid metabolism, further showcasing this miRNA's ability to regulate cholesterol homeostasis. Targeting these metabolic pathways through anti-mi-R33 resulted in improved cholesterol efflux, decreased foam cell formation, and reduced atherosclerotic lesions in atherosclerosis mouse models. The full therapeutic potential of manipulating ncRNAs in the regulation of cardiovascular disease, including cardiac inflammation, remains unclear; however, ncRNAs can be targeted to improve metabolic dysfunction in non-human primates.51 Therefore, ncRNAs could modulate the immunometabolic rewiring of macrophages after myocardial infarction by controlling the expression of metabolic genes. Consequently, manipulation of miR-33 and other relevant ncRNAs may fine-tune the inflammatory response after cardiac injury. ncRNAs may both adjust the regulation of gene expression, which is important during cardiac repair, and further encode human-specific forms not found in experimental rodents.
In macrophage-derived foam cells, both the ncRNA DAPK-IT1 and lipoprotein lipase (LPL) were upregulated after increased LDL exposure.52 In a separate co-expression gene network analysis, DAPK-IT1 was identified as a differentially expressed lncRNA during the progression of atherosclerosis.53 Increased levels of DAPK-IT1 in THP1 macrophages were also associated with reduced expression of miR-590-3p. In this context, in a subsequent analysis of ApoE-deficient mice, DAPK-IT1 was shown to promote LPL expression through the suppression of miR-590-30 in foam cells. These data are consistent with the proposal that the DAPK-IT1/miR-590-3p/LPL axis plays a regulatory role in cholesterol homeostasis and inflammation. Like miR-33, one of the mechanisms through which the DAPK-IT1/miR-590-3p/LPL axis promotes atherogenic effects is through downregulation of the key reverse cholesterol transporters, ABCA1 and ABCG1 (Fig. 1).
The lncRNA growth arrest-specific transcript 5 (GAS5) encodes several functional small nucleolar RNAs that have been shown to act as regulators of apoptosis, as well as a number of other inflammatory conditions such as hepatocellular carcinoma and osteoarthritis. GAS5 has also been identified as a possible biomarker of coronary artery disease54 and cardiac fibrosis.55 In patients with coronary artery disease, GAS5 expression was lower than in healthy controls.54 Like other lncRNAs described in this review, GAS5 also has the capacity to act as a miRNA sponge. In active cardiac fibroblast tissue, GAS5-induced suppression of miR-21 resulted in inhibition of fibrosis.55 These studies suggest that GAS5 may play a protective role in cardiovascular disease. Interestingly, GAS5 appears to have opposite effects in the aggravation of atherosclerosis. In THP1 macrophages, increased expression of GAS5 further increased the ox-LDL-induced release of the proinflammatory cytokines IL-6, IL-1β, and TNF-α.56 Increased expression of GAS5 was also associated with the upregulation of matrix metalloproteinases (MMPs), leading to degradation of the extracellular matrix and atherosclerotic plaque rupture.56 Specifically, GAS5 expression suppressed miR-221, which has been shown to decrease MMP expression and to stabilize atherosclerotic plaques through various anti-inflammatory pathways.56 In a separate study, GAS5 was shown to act as a sponge of miR-135a, another anti-atherosclerotic miRNA.57 Silencing GAS5 led to decreased expression of IL-6, IL-1β, and TNF-α, as depicted in Fig. 1.57 Taken together, these studies suggest that GAS5 plays a pro-inflammatory role in atherosclerosis. It would be interesting to further characterize the role of GAS5 in the development and progression of various CVDs. Research into the role of GAS5 in CVD currently remains limited, and further studies are needed to uncover its full functionality and possible therapeutic potential.
CONCLUSION
Taken together, lncRNAs are a fascinating area of research with implications for many disease processes. In the context of CVD and macrophages, lncRNAs appear to encode a myriad of functions, ranging from transcriptional activation to regulation of immunometabolism and macrophage polarization. This also includes the potential to regulate the clearance of dying cells by the lncRNA MIAT.58 Efferocytosis is critical to the resolution of atherosclerosis and myocardial infarction.59 Significant insights have been gleaned connecting lncRNAs with the regulation of cholesterol homeostasis and miRNAs in macrophages. Future research is necessary to shed light on the full therapeutic potential of targeting ncRNAs relative to other standards of care.
 XML Download
XML Download