

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Endothelial cells (ECs) are cells lining the luminal surface of blood and lymphatic vessels, and ECs in direct contact with blood are referred to as vascular ECs [1]. Loss or dysfunction of vascular ECs usually results in vascular diseases such as atherosclerosis, which can seriously affect human health [2]. Human induced PSCs (iPSCs) are a promising cell source for auto-transplantation when treating vascular diseases [3] and have properties similar to those of embryonic stem cells (ESCs) [45]. They can maintain rapid proliferation and differentiate into other types cells. Patient-specific ECs can be used for drug screening or studying endothelial dysfunction mechanisms [6]. The use of patient-derived iPSCs in transplantation therapy avoids immune rejection and ethical issues [7].
The differentiation of human and mouse iPSCs into ECs has been widely reported, and researchers have investigated ways to optimize methods for promoting cell differentiation and purifying derived cells [89]. A recent study on human cells indicated that ECs with high differentiation efficiency can be cultured [8]. Those authors cultured human ESCs and iPSCs with GSK3i for 2 days, followed by bone morphogenetic protein 4 (BMP4), fibroblast growth factor 2 (FGF2), and vascular endothelial growth factor (VEGF) treatment for 2 days. After 3 days of cultivation with endothelial cell growth medium-2 (EGM-2) and one subculture event, ECs with high purity were obtained. Their study provided an efficient protocol for the differentiation of ECs from human pluripotent cells.
Although differentiation of autologous ECs is promising, clinical application of ECs requires further preclinical exploration and validation in animal models [10]. Typically, the pig is considered an ideal large-animal model of humans, because they are similar in size, have physiological characteristics similar to those in human organs, and have a relatively long-life span [11]. Thus far, many groups have established porcine iPSCs (piPSCs) by applying various methods [12131415], but few have obtained ECs from piPSCs. Differentiation of piPSCs into ECs can facilitate the study of human cardiovascular disease in a porcine model. In 2013, Gu et al. [16] derived ECs from piPSCs via embryoid body differentiation. However, they used several growth factors and chemical compounds, and their protocol was complex and time consuming. In addition to such iPSC origins, some researchers have derived ECs from porcine adipose-derived mesenchymal stem cells (MSCs) and bone marrow MSCs [171819]. Although MSCs are characterized by their ability to differentiate into a wide range of mesodermal cell types, they cannot support long-term in vitro passage.
In this study, we established piPSCs from porcine embryonic fibroblasts (PEFs) and differentiated the cells into ECs. The porcine iPS-ECs expressed endothelial markers and showed comparable morphological and functional properties to immortalized porcine aortic endothelial cells (AOCs). Our study has provided an optimized single-layer culture method for obtaining ECs from porcine PSCs, a method that can be helpful in the study of cardiovascular disease in a pig disease model.
MATERIALS AND METHODS
Ethics statement
All experiments involving animals were approved and conducted according to the guidelines of the Laboratory Animal Ethics Committee of Northeast Agricultural University, P. R. China. The approval number is NEAUEC20160101. We performed all surgeries under anesthesia using isoflurane and tried our best to minimize animals suffering.
Cell culture
PEFs derived from 33.5-day-old embryos were cultured in high-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco, USA) containing 1% penicillin-streptomycin (Gibco), 1% nonessential amino acids (Gibco) and 15% fetal bovine serum (FBS; Hyclone, USA). Mouse embryonic fibroblasts (MEFs) were obtained from 13.5-day-old embryos and were treated with 10 μg/mL mitomycin C (Sigma, USA) as a feeder layer. The piPSCs were cultured in MX medium containing 10% knockout serum replacement (Gibco), 1% penicillin-streptomycin, 0.5% nonessential amino acids, 1mM L-glutamine (Sigma), 0.25% N2 (Gibco), 0.5% B27 (Gibco), 0.25 mg/mL bovine serum albumin (BSA; Sigma), 8 ng/mL basic FGF2 (R&D Systems, USA), 1,000 U/mL human leukemia inhibitory factor (LIF; Millipore, USA), 24% DMEM/F12 (Gibco), 24% Neurobasal (Gibco), and 38% knockout DMEM (Invitrogen, USA). The immortalized porcine AOC cell line was purchased from Abmgood (Canada) and the cells were cultured EGM-2 (Lonza, USA).
Generation of piPSCs
PMX plasmids containing mouse octamer-binding transcription factor 4 (Oct4), sex determining region Y-box (Sox) 2, Kruppel like factor 4 (Klf4), c-Myc, and vesicular stomatitis virus G (VSV-G) were purchased from Addgene (USA). The GP2-293 cells were transfected with 21 μL PLUS and 42 μL LTX reagent (Invitrogen). The pMXs-vector (21 μg) and VSV-G were co-transfected into GP2-293 cells at a ratio of 16:5 in Opti-MEM medium (Gibco). The medium was replaced with DMEM containing 2% FBS after 12 h. Supernatant containing virus was collected at 36 h and 54 h after transduction, and filtered through a 0.45 μm filter. The supernatant was concentrated by centrifugation at 12,000 ×g in a centrifugal filter unit (Millipore). About 1 × 104 PEFs per well were infected with the 4 retroviruses for 24 h in the presence of 4 μg/mL polybrene (Sigma). After 2–3 days, the infected PEFs were passaged at a ratio of 1:3 to MEF feeder cells and cultured in MX medium for another 4–6 days. The MX medium was replaced every day. ESC-like colonies were picked at day 7–8 following a standard protocol.
Differentiation of ECs from piPSCs
Before differentiation, piPSCs were passaged twice on Matrigel (1:100 to 1:150) (Corning, USA) to preclude the presence of MEFs. The piPSCs were then digested by 0.5 μM ethylenediaminetetraacetic acid (EDTA; Thermo, USA) and replanted on Matrigel at dilutions of 1:10 to 1:15 with 10 μM ROCK inhibitor Y27632 (EMD4 Biosciences, USA). MX medium was used and was replaced every day. For differentiation, FGF2 and LIF were removed from the medium (differentiation medium). To explore the effects of CHIR99201 and BMP4 on lineage differentiation during EC derivation, 6 groups were treated as indicated (Table 1). Briefly, the 6 groups were: group F (day 0–2: differentiation medium, day 3–4: differentiation medium with 50 ng/mL FGF2); group FB (day 0–2: differentiation medium, day 3–4: differentiation medium with 50 ng/mL FGF2 and 25 ng/mL BMP4); group BFB (day 0–2: differentiation medium with 25 ng/mL BMP4, day 3–4: differentiation medium with 50 ng/mL FGF2 and 25 ng/mL BMP4); group CF (day 0–2: differentiation medium with 5 μM combination of GSK3β inhibitor [CHIR99021], day 3–4: differentiation medium with 50 ng/mL FGF2); group CFB (day 0–2: differentiation medium with 5 μM CHIR99021, day 3–4: differentiation medium with 50 ng/mL FGF2 and 25 ng/mL BMP4); group CBFB (day 0–2: differentiation medium with 5 μM CHIR99021 and 25 ng/mL BMP4, day 3–4: differentiation medium with 50 ng/mL FGF2 and 25 ng/mL BMP4). From day 5 to day 7, all 6 groups were cultured in differentiation medium with 50 ng/mL VEGF (Novoprotein, USA) and 25 ng/mL BMP4 (R&D Systems). The cells were passaged at a ratio of 1:20 and plated on Matrigel (1:100 to 1:150) with 10 μmol/mL Y27632 in EGM-2 at day 7. On day 10 of differentiation, piPS-ECs were dissociated with 0.05% Trypsin-EDTA and sorted using FACSCalibur (BD Biosciences, USA) and porcine cluster of differentiation (CD) 31 488-conjugated antibody. The separated cells were cultured on Matrigel in EGM-2, which was replaced daily.
Table 1
Different treatments from day 0 to day 4 in 6 different groups

| Group | 0–2 days | 3–4 days |
|---|---|---|
| F | Null | FGF2 |
| FB | Null | FGF2+BMP4 |
| BFB | BMP4 | FGF2+BMP4 |
| CF | CHIR99021 | FGF2 |
| CFB | CHIR99021 | FGF2+BMP4 |
| CBFB | CHIR99021+BMP4 | FGF2+BMP4 |
BMP4, bone morphogenetic protein 4; CHIR99021, combination of GSK3β inhibitor; FGF2, fibroblast growth factor 2.
![]()
RNA isolation and quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR)
Total cellular messenger RNA was extracted using a Pure Link RNA mini kit (Life, USA). The RNA was reverse transcribed to complementary DNA (cDNA) using a High Capacity cDNA Reverse Transcription Kit (ABI, USA). The cDNA was used for qRT-PCR in the presence of specific primers and SYBR Green PCR Master Mix (Takara, USA) [14]. Quantitative PCR was performed on a 7900HT sequence detection system (ABI). Primers for the genes are listed in Supplementary Table 1.
Immunofluorescence staining
The cells were fixed in 4% paraformaldehyde for 30 min at room temperature and washed 3 times in phosphate-buffered saline (PBS). They were then permeabilized in 1% Triton X-100 for 1 h at 37°C followed by blocking with 1% BSA. The cells were then incubated with primary antibodies overnight at 4°C. After washing 3 times with PBS, the cells were incubated with secondary antibodies for 1 h at 37°C. Nuclei were stained with Hoechst 33342, and the labeled cells were visualized on a Nikon 80i fluorescence microscope and images were captured with a Hamamatsu CCD camera. The antibodies are listed in Supplementary Table 2.
Karyotype analysis
The piPSCs were treated with 0.1 μg/mL colchicine for 2.5 h to induce metaphase arrest. The cells were then digested enzymatically to form a single cell suspension in 0.25% trypsin-EDTA and centrifuged at 800 r/min for 10 min. The obtained piPSCs were resuspended with a hypotonic solution (0.1 M KCl medium pre-warmed to 37°C) for 15 min. After another centrifugation at 800 r/min for 10 min, the supernatant was discarded, and a pre-chilled fixative solution (methane:acetic acid = 3:1) was added to resuspend the cells. Tubes with cells were held at 4°C for 30 min and then centrifuged at 800 r/min for 10 min. This process was performed 3 times, and the piPSCs were finally resuspended in 0.4 mL of cold fixative solution. The piPSCs were then dripped onto a slide and stained with 10 mL Giemsa stain. The chromosomes of piPSCs were observed under a Nikon 80i fluorescence microscope.
Teratoma formation
For teratoma formation, the piPSCs were cultured with MX medium on MEF feeder layers for 4 days to achieve 95% confluence. Approximately 1 × 106 piPSCs were resuspended in PBS and injected under the skin of non-obese diabetic/severe combined immune-deficient (NOD/SCID) mice. The injected piPSCs formed tumors within 4 weeks and the teratomas were harvested for histological analysis.
Histological analysis
After the teratomas were collected, they were fixed in a 10% formalin solution for 12 h. The teratomas were then exposed to different concentrations of alcohol for dehydration before being exposed to xylene for 2 h and infiltrated with paraffin for 2 h. The teratomas were embedded into a paraffin wax block to enable sectioning. Obtained paraffin sections were stained with hematoxylin and eosin and observed under a Nikon 80i microscope [20].
Flow cytometric analysis
On day 10 after EC differentiation, the adherent cells were harvested using 0.05% trypsin-EDTA and resuspended in PBS. The cells were then counted and prepared for antibody staining. The anti-porcine CD31 antibody (porcine CD31/PECAM-1 Alexa Fluor 488-conjugated antibody from R&D Systems) was prepared according to the manufacturer's protocol. The cells were incubated with the antibody for 30 min at room temperature, washed 3 times in PBS, and centrifugated at 300 ×g for 5 min. After processing, the labeled cells were sorted by performing flow cytometry on a FACSCalibur cell sorter (BD Biosciences) and analyzed with Flowjo software (BD Biosciences).
In vitro capillary network formation assay
For in vitro capillary network formation, 96-well plates (Corning) were incubated with 60 μL Matrigel (Matrigel Basement Membrane Growth Factor Reduced Matrix, Phenol Red Free; Corning) at 37°C for 30 min, after which, the cells were plated at a density of 1.3 × 104 cells per well in pre-treated plates in EGM-2. Images were taken after 6 h of cultivation.
Low-density lipoprotein (LDL) uptake assay
To identify the ability of the iPS-ECs to phagocytize LDL, the cells were incubated with 20 μg/mL DiI-Ac-LDL (Human DiI-Acetylated LDL; YEASEN, China) in EGM-2 for 4 h at 37°C. The cells were then washed with PBS several times and the images were obtained by using a Nikon80i camera.
Statistical analysis
All experiments were performed at least 3 times unless otherwise stated. All statistical analysis was completed using GraphPad Prism5 software (GraphPad Software, USA). Multiple comparison tests were performed by applying ANOVA with Bonferroni's post-hoc test and determining 95% confidence intervals. Data are shown as mean ± standard deviation values and p < 0.05 was considered significant.
RESULTS
Generation of piPSCs
PEFs were infected by 4 mouse retroviral reprogramming factors: Sox2, Klf4, Oct4, and c-Myc (Fig. 1A). The initial time of infection was designated as day 0. The infected cells were plated on MEF feeder layer cells in MX medium on day 3. Approximately 10 to 11 days after viral transduction, iPSC-like colonies emerged. The colonies were dissociated mechanically and passaged using type IV collagenase.
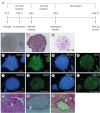 | Fig. 1Identification of piPSCs. (A) Schematic diagram of the piPSC generation procedure. (B) piPSCs (Pilw4, passage 7) in bright field. (C) Alkaline phosphatase staining of piPSCs. (D) Example of karyotype analysis results of piPSCs. (E-L) Immunofluorescence staining results show that piPSCs were positive for Oct4, Sox2, SSEA3, and SSEA4. (M-O) Teratomas derived from piPSCs reveal contribution of the cells to 3 germ layers. Nervous tissues representing ectoderm (M); cartilage tissue representing mesoderm (N); intestinal gland tissues representing endoderm (O). Scale bar = 100 μm.FBS, fetal bovine serum; piPSC, porcine induced pluripotent stem cell; Oct4, octamer-binding transcription factor 4; Sox2, sex determining region Y-box 2; SSEA, stage-specific embryonic antigen.
|
Five piPSC lines were established (Pilw1–Pilw5), and 3 of them (Pilw2, Pilw4, and Pilw5) were used for endothelial differentiation. The piPSCs appeared flattened and were alkaline phosphatase stain-positive (Fig. 1B and C). The cell lines could be maintained in vitro for more than 30 passages and each exhibited a normal karyotype of 38 chromosomes (Fig. 1D). Immunofluorescence analysis indicated that the piPSCs expressed the pluripotent genes Oct4, Sox2, stage-specific embryonic antigen (SSEA) 3, and SSEA4 (Fig. 1E–L). Moreover, the piPSCs formed teratomas that contained tissues from 3 germ layers after injection into NOD/SCID mice (Fig. 1M–O).
Expression of lineage-specific genes under different treatments during endothelial differentiation from piPSCs
Before differentiation, piPSCs were maintained on Matrigel-coated plates in MX medium and routinely passaged every 4 days. When the density of cells was approximately 90%, the MX medium was exchanged with differentiation medium. During EC differentiation, different combinations of small-molecule inhibitors and cytokines were applied to the cells (Fig. 2 and Table 1). At day 10, CD31-positive cells were isolated by flow cytometry assisted sorting, and plated on 0.1% gelatin-coated plates in the presence of EGM-2.
 | Fig. 2Flow chart for porcine induced pluripotent stem cell-derived endothelial cells derivation.FGF2, fibroblast growth factor 2; LIF, leukemia inhibitory factor; EGM-2, endothelial cell growth medium-2; BMP4, bone morphogenetic protein 4; CHIR99021, combination of GSK3β inhibitor; VEGF, vascular endothelial growth factor; FACS, fluorescence-activated cell sorting; CD, cluster of differentiation.
|
During endothelial differentiation, there were no morphological or quantitative changes of the cells in the F, FB, or BFB groups, whereas, in the other 3 groups, cell quantity decreased during the first 2 days, especially in the CBFB group. After 7 days of cultivation in Matrigel-coated plates, some flat epithelium-like cells emerged in the CFB and CBFB groups. We analyzed expression levels of some lineage-specific genes such as primitive streak markers, mesoderm markers, and endoderm markers during day 0–4 of differentiation using qRT-PCR. Among primitive streak markers, we observed increases in Brachyury and MIXL1 at day 1 and 2 in CF, CFB, and CBFB groups, which subsequently decreased; however, there were no obvious changes in those markers in the F, FB, and BFB groups (Fig. 3A and B, Supplementary Table 3). The kinase insert domain receptor marker kept increasing during day 0–4 in all 6 groups. Among the mesoderm markers, there were marked increases of mesoderm posterior (Mesp) 1 and Mesp2 at day 3 and of platelet-derived growth factor receptor (PDGFR) α and PDGFRβ at day 4 in the CF, CFB, and CBFB groups, but approximately constant levels were maintained in the other 3 groups (Fig. 3C–G, Supplementary Table 3). Among the endoderm markers, alpha-fetoprotein, hepatocyte nuclear factor 4α, and Sox17 were up-regulated and then down-regulated in the CF, CFB, and CBFB groups, but there were no notable changes in expressions of these marker genes in the other 3 groups (Fig. 3H–J, Supplementary Table 3). Our results indicate that CHIR99021 treatment at the beginning of EC differentiation promotes primitive streak differentiation, while a combination of BMP4 and FGF2 treatment on the following days can promote mesoderm differentiation. In addition, BMP4 treatment combined with CHIR99021 and FGF2 throughout day 0–4 induced the highest expressions among the tested genes.
 | Fig. 3Expression levels of lineage-specific genes during day 0–4 of differentiation. (A-J) During day 0–4 of porcine induced pluripotent stem cell-derived endothelial cells differentiation, expression levels of some lineage-specific genes were detected by quantitative real-time reverse transcription polymerase chain reaction. (A, B) Primitive streak marker genes Brachyury and MIXL1; (C-G) mesoderm marker genes KDR, PDGFRα, PDGFRβ, Mesp1, and Mesp2; and (H-J) endoderm marker genes AFP, HNF4α, and Sox17 were chosen for detection. Three experimental replicates were performed.KDR, kinase insert domain receptor; PDGFR, platelet-derived growth factor receptor; Mesp, mesoderm posterior; AFP, alpha-fetoprotein; HNF4α, hepatocyte nuclear factor 4α; Sox17, sex determining region Y-box 17.
|
Effects of different treatments on EC differentiation from piPSCs
On day 10 of differentiation, the cells in different groups were digested with 0.05% trypsin-EDTA to obtain single cells. Some cells were used to perform qRT-PCR to determine the expression levels of endothelial markers, while others were used to determine the percentages of CD31-positive cells. The expression levels of endothelial markers in the CF, CFB, and CBFB groups were higher than those in the other 3 groups (Fig. 4A; p < 0.05). Moreover, the CFB and CBFB groups had higher expression levels of CD31, CD144, and von Willebrand factor (vWF) than those in the CF group, and their vWF expression levels were comparable to that in AOC cells (Fig. 4A; p < 0.05). Flow cytometric analysis revealed that the CFB group had the highest percentage of CD31-positive cells (20.92% ± 0.58%), while in the groups without a CHIR99021 treatment, there were only 3.35% ± 0.37% to 5.32% ± 0.46% CD31-positive cells (Fig. 4B and C). These results indicate that CHIR99021 has an important role in the initiation of EC differentiation from piPSCs. In addition, we observed that the addition of BMP4 in the middle differentiation stage improved EC differentiation efficiency; for example, the CFB group treated with BMP4 on day 3–4 of differentiation had 20.92% ± 0.58% CD31-positive cells while the percentages were only 12.43% ± 0.33% in CF group and 16.85% ± 0.47% in the CBFB group (Fig. 4B and C). Thus, in the following part of this study, we adopted the optimized culture conditions of the CFB group.
 | Fig. 4Effects of different treatments on EC differentiation from piPSCs. (A) On day 10 of differentiation, expression levels of EC marker genes were analyzed by quantitative real-time reverse transcription polymerase chain reaction. The piPSCs were used as control samples. Three experimental replicates were performed. All data are presented as mean ± standard deviation values. Different letters indicate significant differences (p < 0.05). (B) On day 10 of differentiation, the cells were digested to single cells for immunofluorescence staining of CD31. CD31-positive cells were sorted through FACSCalibur. The piPSCs were used as negative control samples. Representative percentages of CD31-positive cells in F, FB, BFB, CF, CFB, and CBFB groups are shown. (C) Mean percentages of CD31-positive cells in the various groups. Different superscript letters between groups indicate significant differences between groups.EC, endothelial cell; piPSC, porcine induced pluripotent stem cell; CD, cluster of differentiation; KDR, kinase insert domain receptor; vWF, von Willebrand factor.
|
Physiological characterization of the piPS-ECs
Next, we induced piPSCs into ECs using the abovementioned optimized culture conditions. AOCs were used as the positive control (Fig. 5G–J) and piPSCs were used as the negative control (Supplementary Fig. 1). On day 10 of differentiation, CD31-positive cells were obtained by flow cytometric sorting and cultured on gelatin with EGM-2. The morphology of the piPS-ECs was similar to that of porcine AOCs (Fig. 5A and B), and the immunofluorescence staining results showed that the piPS-ECs expressed endothelial markers CD31, CD144, vWF, and endothelial NO synthase (Fig. 5C–F). Since the uptake of LDL is a functional characteristic of ECs, we then undertook to detect uptake by piPS-ECs. The results showed that, similar to AOCs (Fig. 5M and N), piPS-ECs were capable of Dil-Ac-LDL uptake (Fig. 5K and L), but piPSCs were incapable of Dil-Ac-LDL in the (Supplementary Fig. 1). In addition, piPS-ECs could assemble into tube-like structures after 6 h of culturing on Matrigel (Fig. 5O); AOC, the positive control, results are presented in Fig. 5P. Our results showed that the piPS-ECs not only exhibit morphological characteristics and gene expression patterns similar to those of ECs, but also, they possess comparable functions to those of ECs.
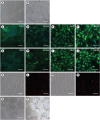 | Fig. 5Characterization of the derived piPS-ECs. (A) Typical microscope images of piPSC-ECs 1 day after flow cytometric sorting; scale bar = 80 μm. (B) Typical microscope images of piPSC-ECs when they reached 90% confluence; scale bar = 80 μm. (C-J) EC-specific markers were detected via immunofluorescence staining. The piPS-ECs were positive for CD31, CD144, vWF, and eNOS (C-F); scale bar = 20 μm. AOCs were positive for CD31, CD144, vWF, and eNOS (G-J); scale bar = 20 μm. (K-N) Uptake of Dil-Ac-LDL y piPSC-ECs (K-L) and AOCs (M-N); scale bar = 20 μm. (O-P) Tube-like structures were formed by piPS-ECs (O) and AOCs (P); scale bar = 80 μm.CD, cluster of differentiation; vWF, von Willebrand factor; eNOS, endothelial NO synthase; EC, endothelial cell; piPS-EC, porcine induced pluripotent stem cell-derived endothelial cell; AOC, aortic endothelial cell; EC, endothelial cell; piPSC, porcine induced pluripotent stem cell.
|
DISCUSSION
In this study, we generated ECs from piPSCs via a feeder-free monolayer culture system and characterized the ECs expression patterns and functional phenotypes. We detected a combination of effects from several growth factors and chemical molecules in the early stage of differentiation and observed that the addition of both CHIR99021 and BMP4 promoted mesodermal differentiation and resulted in a high rate of CD31-positive cell production.
CHIR99021 can inhibit GSK3β, resulting in Wnt signaling activation in mouse ESCs, and it can synergize with the MEK/ERK signaling inhibitor PD325901 to maintain mouse ESC self-renewal. Previous study has shown that short-term treatment of CHIR99021 can initiate primitive streak differentiation in human ESCs [21]. Our study gave a result consistent with that finding as the groups treated with CHIR99021 displayed significantly increased expression of primitive streak markers, while the groups without CHIR99021 treatment exhibited nearly no change in the expression of these markers. Furthermore, we obtained more CD31-positive cells on day 10 of differentiation in the CHIR99021-treated groups than in the untreated ones (Fig. 4B and C), indicating that the GSK3β signaling pathway is important in piPSCs differentiation, similar to that observed in human and mouse cells [2122] in which where the inhibition of GSK3β signaling is critical for piPSC differentiation into ECs.
During embryonic development, BMP4 has an important role in mesoderm differentiation and vascular endothelial progenitor generation [2324]. Knockout of BMP4 leads to early embryonic lethality [4], and knockout of its downstream effectors, such as Smad4 or Smad5, causes failure of mouse embryo mesoderm formation and abnormal visceral endoderm development [525]. In the process of cell differentiation in vitro, the role of BMP4 is not the same in human and mouse ESCs. During mouse ESC differentiation, activation of BMP4 signaling enhances the formation of a mesoderm cell population, resulting in hematopoietic and endothelial progenitor cells [26], while in human ESCs, BMP4 is a direct regulator that initiates EC differentiation [27]. Moreover, previous study has shown that a combined treatment of BMP4 and VEGF results in a higher rate of CD31-positive cell production than that in VEGF treated alone cells [28]. In our study, the expression levels of most mesoderm markers in the CFB and CBFB groups were much higher than those in the CF group after the combined treatment of FGF2 and BMP4 for 3 days (Fig. 3), at which time the expression levels of primitive streak markers sharply declined in all 3 groups. On day 10 of differentiation, the CFB group had the highest expression levels of endothelial markers and the highest proportion of CD31-positive cells (Fig. 4C). Thus, we conjectured that BMP4 promoted mesoderm differentiation of piPSCs and also improved their endothelial differentiation efficiency. Our findings suggest that the role of BMP4 in piPSC differentiation is similar to that in mouse ESC differentiation.
Several methods have been successfully used to induce differentiation of human and mouse ESCs or other PSCs into ECs [9293031], and several researchers are engaged in optimizing cell differentiation and purification methods [9323334], via the study of signaling mechanisms that are key to endothelial formation [83536], and testing the function of derived cells in vivo and in vitro [89]. But there were few reports on EC differentiation in large animals such as pigs. The pig is considered an ideal model because of its size and physiological features [11]. Many researchers have focused on the establishment of piPSCs rather than on the differentiation of piPSCs into ECs [12131415]. In 2013, Gu et al. [16] derived ECs from piPSCs via an embryoid body differentiation method over a period of 12 days and using a complex differentiation medium that contained Activin A, BMP4, FGF2, VEGF, and the transforming growth factor-β inhibitor SB431542. Since those researchers did not study the specific effects of each added factor or compound, lineage differentiation during porcine endothelial derivation remained ambiguous. Other research groups have derived ECs from porcine MSCs in recent years with different differentiation protocols; however, from those different protocols we can at least conclude that VEGF was needed to induce porcine MSCs to differentiate into ECs [1718]. However, compared with PSCs, MSCs have limited pass-through capacity which cannot be passaged in vitro for a long time, thus it is difficult to generate the large amount of cells differentiated from MSCs for clinical transplantation. Moreover, differentiation of human or mouse PSCs into ECs via monolayer culture method is more mature and more efficient (about 60%), so further research is needed to improve the endothelial differentiation efficiency from porcine PSCs and via monolayer culture method.
In this study, using a 2-dimensional feeder-free culture system, we derived functional ECs from piPSCs. In addition, we explored the effects of different combinations of small-molecule inhibitors and cytokines on lineage differentiation during endothelial differentiation and observed that the combined addition of CHIR99021 and BMP4 promoted mesodermal differentiation and resulted in higher rates of EC production. Herein, we describe an optimized single-layer culture method for obtaining ECs from porcine PSCs, which is the first report on such a method that is applicable to a species other than human and mouse.
 XML Download
XML Download