

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis is a prolonged skin disease characterized by allergic inflammation showing erythema, hemorrhage, dryness, edema, erosion, crust, and skin thickening [12]. The prevalence of atopic dermatitis has increased in recent years, and it is now estimated to affect up to 20% of the general population [3]. Although immunosuppression drugs such as adrenocortical steroids, anti-histamines, cyclosporin A, and FK506 can be used to treat chronic dermatitis, they have many side effects including nephrotoxicity, osteoporosis, hypertension, and severe pruritus [456]. In recent years, therefore, there has been considerable interest in natural compound treatments that have no side effects.
Horse oil has been used as a folk medicine for the treatment of burn wound or skin damage in many Asian countries, including Korea, Mongolia, China, and Japan. Based on traditional Chinese medical books, horse oil has healing effects for burn wounds and skin damage [789]. Horse oil is comprised of 60%–65% unsaturated fatty acids, its composition is close to that of sebum produced by sebaceous glands in human beings, and it is easily absorbed by human skin [10]. Horse oil is reported to produce anti-inflammatory effects by reducing erythema and IgE level and recovering histopathological signs such as thickening of epidermis, hyperkeratosis, and infiltration of inflammatory cells in 2, 4-dinitrochlorobenzene (DNCB)-treated BALB/c mice [10].
Despite various effects of horse oil being described in many Asian countries, scientific evaluation of those effects is incomplete. The objective of this study was to investigate the effect of horse oil on DNCB-induced atopic dermatitis in BALB/c mice and to evaluate its therapeutic potential as a topical application for atopic dermatitis healing. To accomplish this, we induced atopic dermatitis by applying DNCB on the dorsal skin of BALB/c mice. After the topical application of horse oil, the pathological features of the skin were examined, and the differential gene expression profiles of the treatment groups were observed by undertaking cDNA microarray analysis.
MATERIALS AND METHODS
Animal experiments
The study was performed using male BALB/c (8-week-old) mice purchased from Orient Bio (Korea) and kept in a storage room under a constant temperature of 24°C ± 1°C, relative humidity of 50%–55%, and illumination provided in 12 h light/dark cycles. All animal experiments were carried out in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee of Jeju National University (Approval 2013-0017). The mice were allocated into 4 groups of 5 animals each; Group 1 was treated with distilled water only. Group 2 was treated with DNCB (Sigma Aldrich, USA). Group 3 was treated with DNCB and Atobarrier lotion (AB; Amore Pacific Co., Korea). Group 4 was treated with DNCB and horse oil lotion (Jeju Mayu Co., Korea). The mice in groups 2 to 4 were treated with 100 μL of 1% DNCB on their dorsal skin twice, 3 days apart, for sensitization and were similarly treated with 100 μL of 0.2% DNCB twice, 3 days apart, as challenge exposures. After the second challenge exposure, groups 3 and 4 received topical treatments of AB or horse oil lotion (100 mg), respectively, on their dorsal skin. The topical treatments were performed once a day for 7 consecutive days. After inducing atopic dermatitis with DNCB in BALB/c mice, the clinical signs of erythema, hemorrhage, dryness, edema, erosion, crust, and skin thickening were observed. After the end of the treatments, the mice were killed by CO2 application, and a sample of the dorsal skin (1 cm × 1 cm) was removed for RNA preparation.
RNA preparation
Total RNA was isolated from mouse skin samples (1 cm × 1 cm) using an Easy-blue RNA extraction kit (Intron, Korea) according to the manufacturer's recommended protocol. The amount and purity of each RNA sample were determined by spectrophotometry at 260/280 nm. Quality of total RNA was based on the migration pattern in electrophoresis, the ratio of 28s/18s ribosomal RNA, and the RNA integrity number (RIN) obtained by using the Agilent 2100 Bioanalyzer system (Agilent Technology, USA). A 28s/18s ribosomal RNA ratio higher than 1.5 and a RIN value higher than 7.5 were used for the analysis.
cDNA microarray analysis
Transcription of double-stranded DNA was performed by adding the transcription master mix to the double-stranded DNA reaction samples, after which the mixture was incubated at 40°C for 2 h. Amplified and labeled cRNA was purified using cRNA Cleanup Module (Agilent Technology) according to the manufacturer's protocol. Labeled target cRNA was quantified using an ND-1000 spectrophotometer (NanoDrop Technologies, USA). After determining the labeling efficiency, fragmentation of cRNA was performed by adding 10× blocking agent and 25× fragmentation buffer followed by incubation at 60°C for 30 min. The fragmented cRNA was resuspended with 2× hybridization buffer and directly pipetted onto assembled Mouse Oligo Microarray 44K (Agilent Technology). The arrays were hybridized at 65°C for 17 h using an Agilent hybridization oven (Agilent Technology). The hybridized microarrays were washed per the manufacturer's washing protocol (Agilent Technology). The hybridized images were scanned using Agilent's DNA microarray scanner (Agilent Technology) and were quantified by using Agilent Feature Extraction Software (Agilent Technology). Functional annotations of genes were performed in accordance with the protocols of the Gene Ontology Consortium [11] by using GeneSpring GX 7.3 software (Agilent Technology). Gene classification was based on searches using MEDLINE databases (National Center for Biotechnology Information, USA).
Pathway analysis
Pathway analysis of the differentially expressed genes was established by using pathway data within the Kyoto Encyclopedia of Genes and Genomes database. The level of protein-protein interaction (PPI) was analyzed and graphically visualized as nodes (proteins) and edges (the relationship between proteins) by using Ingenuity Pathway Analysis (IPA; QIAGEN, USA).
RESULTS
The healing effect of horse oil was observed in DNCB-induced atopic dermatitis in BALB/c mice (Fig. 1). After the application of DNCB, the treated mice exhibited more atopic dermatitis symptoms including severe erythema, hemorrhage, and erosion (Fig. 1B) than those shown by the control group (Fig. 1A). In contrast, DNCB-induced symptoms were alleviated after cotreatment with AB (Fig. 1C) or horse oil (Fig. 1D), indicating that both AB and horse oil can reverse the effects of DNCB treatment.
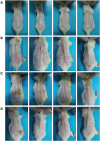 | Fig. 1The anti-atopic dermatitis effect of horse oil in DNCB-treated BALB/c mice. Skin lesions in the (A) control, (B) DNCB, (C) DNCB + AB, and (D) DNCB + horse oil treatment groups are shown.DNCB, 2, 4-dinitrochlorobenzene; AB, Atobarrier lotion.
|
Molecular-level gene expressions in mouse dorsal skin samples were analyzed using cDNA microarray. Total RNA was isolated from the dorsal skins of each group for use in the cDNA microarray analysis. Fig. 2 shows the migration and peak patterns of the isolated RNAs. For each sample, the 28/18 ribosomal RNA ratio was greater than 1.5, and the RIN was above 7.0, indicating that the quality of the total RNA was sufficiently high for use in cDNA microarray analysis. After cRNA labeling, samples were hybridized onto mouse oligo microarray slides, which were then scanned and analyzed (Fig. 3).
 | Fig. 2Isolation of RNAs from the skin of mice. RNAs were isolated from the skin of control-, DNCB-, DNCB + AB-, and DNCB + horse oil-treated mice. (A) Migration pattern (electrophoretic trace), and (B) peak pattern (electropherogram) of RNAs are shown.DNCB, 2, 4-dinitrochlorobenzene; AB, Atobarrier lotion.
|
 | Fig. 3Scanned image of cDNA microarray chips. After labeling of cRNA, samples from the control-, DNCB-, DNCB + AB-, and DNCB + horse oil-treated mice were hybridized onto mouse oligo microarray slides, which were then scanned and analyzed.DNCB, 2, 4-dinitrochlorobenzene; AB, Atobarrier lotion.
|
Based on the cDNA microarray results, among the 39,429 genes in a mouse, 3,089 genes were up-regulated (higher than 2.0-fold) and 393 genes were down-regulated (lower than 0.5-fold) in the DNCB-treated group compared to the control group levels (Table 1). In addition, 2,060 genes were up-regulated and 1,393 genes were down-regulated in the DNCB + AB group compared to the control group levels (Table 2). Moreover, 2,494 genes were up-regulated and 2,763 genes were down-regulated in the DNCB + horse oil group compared to the control group levels (Table 3).
Table 1
Differential expression of functional genes in mice skin treated with dinitrochlorobenzene only

AP, apoptosis process; CC, cell cycle; CD, cell death; CDi, cell differentiation; CM, cell migration; CP, cell proliferation; EM, extracellular matrix; IR, immune response; InR, inflammatory response; NG, neurogenesis.
![]()
Table 2
Differential expression of functional genes in mice skin treated with dinitrochlorobenzene and Atobarrier

AP, apoptosis process; CC, cell cycle; CD, cell death; CDi, cell differentiation; CM, cell migration; CP, cell proliferation; EM, extracellular matrix; IR, immune response; InR, inflammatory response; NG, neurogenesis.
![]()
Table 3
Differential expression of functional genes in mice skin treated with dinitrochlorobenzene and horse-oil

AP, apoptosis process; CC, cell cycle; CD, cell death; CDi, cell differentiation; CM, cell migration; CP, cell proliferation; EM, extracellular matrix; IR, immune response; InR, inflammatory response; NG, neurogenesis.
![]()
Functional annotation of genes was performed based on gene ontology analysis. As shown in Fig. 4, a large portion of the genes related to inflammation and immune response was significantly changed in both the DNCB and DNCB + horse oil groups compared to the control group. Of the genes associated with inflammation, 17% were up-regulated and 6.4% were down-regulated in the DNCB group compared to the control group levels (Fig. 4A). In the DNCB + AB group, 21.8% of the genes related to inflammation were up-regulated and 6.4% of genes were down-regulated compared to the control group levels (Fig. 4B). In the DNCB + horse oil group, however, only 8.5% of genes related to inflammation were up-regulated and 14.7% were down-regulated compared to control group levels (Fig. 4C). The results indicate that inflammatory gene expression levels are changed differently following treatment with DNCB or horse oil.
 | Fig. 4GO analysis of DNCB-treated mouse skin applied with/without AB or horse oil as a percentage of the total significant. The transcripts of the DNCB-treated mouse skin applied with/without AB or horse oil were classified into GO categories of (A) DNCB/control, (B) DNCB + AB/control, and (C) DNCB + horse oil/control based on their GO terms as a percentage of the total significant.GO, gene ontology; DNCB, 2, 4-dinitrochlorobenzene; AB, Atobarrier lotion.
|
The cDNA microarray analysis revealed that the expression levels of 30 genes related to inflammation were up-regulated (higher than 2.0-fold) in the DNCB group compared to the control group. In particular, the gene expression levels of Ccr1, Ccr2, Ccl20, Anxa1, and Hc genes were up-regulated in the DNCB group compared to the control group levels, whereas the levels in the DNCB + horse oil group were similar to those in the control group (Table 4). In contrast, the gene expression levels of 28 genes related to inflammation were down-regulated (lower than 0.5-fold) in the DNCB group compared to the control group levels (Table 5). In particular, the expression levels of several chemokine genes including Ccl5, Ccl7, Ccl8, Cxcl10, and Cxcl13 were down-regulated in the DNCB group compared to the control group levels, whereas those levels were similar to the control levels in the DNCB + horse oil group (Table 5).
Table 4
Up-regulated inflammatory genes in the skin of DNCB-treated mice and normalized by horse-oil

![]()
Table 5
Down-regulated inflammatory genes in the skin of DNCB-treated mice and normalized by horse-oil

![]()
To investigate the relationships among the differentially expressed genes, PPI analysis was performed using STRING analysis. The protein-protein network was visualized as a set of nodes and edges, as shown in Fig. 5. Among the proteins related to inflammation, chemokine ligands (Ccl5, Ccl8, Ccl20, Cxcl10, and Cxcl113) and chemokine receptors (Ccr1, Ccr2, and Ccr3) were closely located and formed a large “functional cluster” in the middle of the network (Fig. 5). Fig. 6 summarizes the relationships among the chemokine ligands and receptors which were up- or down-regulated by DNCB and approximately restored to control levels by horse oil treatment. Overall, the results show that horse oil restores the DNCB-perturbed expression levels of genes related to chemokines to control levels.
 | Fig. 5PPI network constructed for DEGs identified in mouse skin treated with DNCB and horse oil. Representative image of the PPI network is shown. Bold lines represent the strongly connected genes associated with inflammation. Different colors indicate DEGs; red indicates up-regulated gene expression and blue indicates down-regulated gene expression.PPI, protein-protein interaction; DEG, differentially expressed gene; DNCB, 2, 4-dinitrochlorobenzene.
|
DISCUSSION
The present study was conducted to evaluate the anti-inflammatory effect of horse oil in DNCB-treated BALB/c mice. DNCB, an electrophilic and cytotoxic benzene derivative, was used to induce stable clinical atopic dermatitis. Skin changes in DNCB-treated mice were evident by scratching behavior, followed by the rapid development of erythema, edema, and hemorrhage [10]. A previous study reported anti-inflammatory effects of horse oil, which included the alleviation of DNCB-induced histopathological signs such as thickening of the epidermis, hyperkeratosis, and infiltration of inflammatory cells [10]. The present study confirmed that horse oil can reduce atopic dermatitis symptoms including severe erythema, hemorrhage, and erosion in DNCB-treated BALB/c mice.
The results of the present study show that the expression levels of genes related to the inflammation, including Ccr1, Ccr2, Ccl20, Anxa1, and Hc genes, were up-regulated from the control level in DNCB-treated mice, whereas the expression levels were similar to control levels in the DNCB + horse oil-treated mice. Ccr1 induces interstitial inflammation and fibrosis [12], and the Ccr2 gene is reported to promote colonic inflammation [13]. Ccl20 is reported to induce severe inflammation in pediatric ulcerative colitis [14]. In addition, Anxa1 is reported to modulate systemic anti-inflammatory processes [15], which Hc functions in multiple biological processes, including the inflammatory process [2].
In addition, the current study showed that the gene expression levels related to inflammation, including Ccl5, Ccl7, Ccl8, Cxcl10, and Cxcl13 genes, were down-regulated in DNCB-treated mice compared to the levels in control mice, whereas the levels were similar to the control levels in the DNCB+horse oil-treated mice. Ccl5 presents discrete tissue microenvironments during renal inflammation [16], whereas Ccr7 is involved in inflammatory responses to common allergens such as house dust, pollens, and air pollutants [17]. Ccl8, which is secreted by macrophages, is reported to initiate mucosal inflammation through the recruitment of inflammatory monocytes [18]. Cxcl10 is a pro-inflammatory cytokine that activated immune cells and stimulates their migration of immune cells to infected sites [19], whereas Cxcl13 promotes and exaggerates inflammation, actions that are related to B cells and Th cells [1]. The results in the present study suggest that horse oil treatment can restore the DNCB-perturbed expression levels of genes related to chemokines.
Among the proteins related to inflammation, chemokine ligands (Ccl5, Ccl8, Ccl20, Cxcl10, and Cxcl113) and chemokine receptors (Ccr1, Ccr2, and Ccr3) were shown to be closely located, forming a large “functional cluster” in the middle of the protein-PPI network. That result suggests that the chemokine proteins within the “functional cluster” have key roles in the induction of atopic dermatitis by DNCB. Interestingly, as shown in Fig. 6, none of the chemokine ligand-receptor pairs was up- or down-regulated simultaneously by DNCB. We hypothesize that more chemokine receptors are expressed to compensate for the down-regulation of the chemokine ligands, and vice versa, in order to produce homeostasis in the biological system. Further studies are required to evaluate this suggestion.
In the current research, the significance of the observed horse oil effect is related to its ability to restore, to control levels, the expression levels of genes associated with inflammation, that had been up- or down-regulated in BALB/c mice with DNCB-induced atopic dermatitis. In particular, the expression levels of chemokine genes, including chemokine ligands and chemokine receptors, were up- or down-regulated by DNCB, and those levels were subsequently restored to control levels by horse oil treatment. Based on those results, horse oil can be considered as a therapeutic agent for the treatment of skin damaged by atopic dermatitis. In addition, the results observed in the present study may provide valuable information for use in the development of new anti-dermatitis agents. Further studies are necessary to extrapolate the effectiveness of this topical application in mice to that for dermatitis repair in human skin.
 XML Download
XML Download