

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Focal cerebral ischemia (fCI) is one of the leading causes of death and permanent disability worldwide, resulting in irreversible neuronal damage and loss of sensorimotor function [12]. The only clinically available drug directed at fCI is intravenous administration of recombinant tissue plasminogen activator (rT-PA), a thrombolytic agent [3]. When delivered up to 3.5 hours after the initial insult, rT-PA can attenuate fCI-induced brain damage. However, a majority of patients are not admitted to the clinic within this “golden window” after the onset of fCI; thus, treatment with rT-PA often has decreased efficacy [4]. To overcome this, a thousand pharmaceutical candidates have been screened and suggested as the alternatives to rT-PA. However, almost all the trials have been unsuccessful [567]. The resulting pessimism has turned researchers' attention towards the development of a “preventive agent” in an effort to minimize the neuronal damage during a future fCI insult [8910].
Among the preventive candidates, phytochemicals originating from plants are currently attracting substantial attention. Recent studies have revealed that various polyphenols enriched in plants have various health-promoting effects [111213]. They are known to possess significant antioxidant capacity, which is considered the main mechanism of conferring the health benefits [14151617]. Since an imbalance between the generation and the clearance of reactive oxygen species (ROS) is known to be the key event in fCI pathogenesis, it is logical that antioxidant capacity of plant polyphenols may confer protection against fCI [18].
Based on the above assumption, this study tested the protective effects of Platycarya strobilacea (PS) leaf extract (PSL) on fCI pathogenesis. PS is a small deciduous broad-leaf tree belonging to the wild walnut family and is abundant in Asian countries, including Korea [1920]. In traditional medicine, PSL has long been used empirically without scientific rationale. To date, unfortunately, information about the possible efficacies of PSL on specific disease models is rare, except in cases citing its antifungal [21] and anticancer [22] activities. However, recent studies reporting potent antioxidative effects of juglone (5-hydroxy-2-methoxy-1,4-naphthoquinone) [23] and ellagic acid [24], the two key polyphenolic constituents of PSL, suggest that the crude form of PSL may exert a neuroprotective effect. However, there have been no studies elucidating the neuroprotective role of PSL against fCI.
Therefore, this study was designed to investigate the therapeutic effect and the possible underlying mechanism of PSL against fCI pathogenesis. The in vivo therapeutic effect of PSL was examined by measuring infarct volume and the severity of motor deficits, and the antioxidant mechanisms were assessed using oxidative damage-associated apoptosis assays.
Materials and Methods
Preparation of PSL extract
PSL extract was kindly supplied by Lifetree Biotech Co., Ltd. (Suwon, Korea). For the preparation, the leaves of the PS were harvested at Wonju, Gangwon-do, South Korea. The harvested leaves were mixed with water (1:10, w/v), and then eluted with boiling water for 3 hours. The extract was concentrated to a 15 % solid content yield using a vacuum evaporator. The concentrated extract was diluted with an equal volume of dextrin, and freeze-dried in a lyophilizer system (Advantage 2.0, SP Scientific, Warminster, PA, USA). The yield was approximately 4%.
Animals and experimental design
The animal protocol used in this study was reviewed and approved on the basis of the ethical procedures and scientific care by the Institutional Animal Care and Use Committee (IACUC) in Konyang University (Daejeon, Korea). All experimental procedures were performed in accordance with the National Institutes of Health (NIH, Bethesda, MD, USA) Guidelines for the Care and Use of Laboratory Animals, eighth edition [25]. Forty-eight male C57/BL6 mice (25–30 g; 8 weeks) were purchased from Samtako (Osan, Korea). On arrival, the mice were stabilized for 7 days, with access to water and food ad libitum at a constant temperature (22℃±2℃) and humidity (40%–60%) with a 12-hour light/dark cycle. Following this, the mice were randomly divided into four groups as follows (n=12 per group): SO, subjected to a shamoperation; VEH, treated with distilled water as a vehicle and a subsequent operation; PSL-L, treated with low dose (20 mg/kg) of PSL and subjected to an operation; and PSL-H, treated with high dose (100 mg/kg) of PSL and subjected to an operation. The operation technique involved 60 minutes of middle cerebral artery occlusion and reperfusion (MCAO/R). Either vehicle or PSL was administered daily by an intraoral route for 8 days prior to MCAO/R. The operated mice were returned to their home cage and kept until sacrifice. The experimental plan is schematically illustrated in Fig. 1.
MCAO/R
Mice were anesthetized with an intraperitoneal injection of 30 mg/kg ketamine and 20 mg/kg xylazine cocktail. Following a midline neck incision, the left common carotid artery (CCA) and the external carotid artery were ligated. A clip was placed on the internal carotid artery (ICA) to prevent bleeding. After creating a small hole in the CCA, 7-0 nylon monofilament with a round silicon-coated tip (0.2 mm in diameter) was inserted into the hole. Following removal of the clip at the ICA, the silicon-coated filament was further inserted into the ICA to occlude the middle cerebral artery (MCA) until the cerebral blood flow (CBF) was abruptly dropped. When CBF dropped below 20% of the baseline, insertion of the filament was stopped and 60 minutes of occlusion began. After 60 minutes, the filaments were withdrawn and reperfusion began. During the operation, CBF and rectal temperature (37℃±0.5℃) were carefully monitored using a Doppler flowmeter (Periflux 5000, Perimed AB, Stockholm, Sweden) and heating pad, respectively. Sham-operated mice underwent the same surgical procedure, except that the filament was not advanced far enough to occlude the MCA.
In vivo ROS detection assay
To detect brain cellular ROS accumulation we used dihydroethidium (DHE), which is a fluorescent ROS probe. For this procedure, 10 mg/kg of DHE (Invitrogen, Carlsbad, CA, USA) was diluted in 50 µL of normal saline and injected into the mice (n=2 per each group) via a jugular vein just before MCAO/R. At 2 hours after MCAO/R, the brain hemisphere ipsilateral to fCI lesion was removed, immersed in 30% sucrose solution for cryopreservation, embedded in an OCT compound, and frozen in dry ice. Using a cryostat microtome (HM430, Leica, Walldorf, Germany) set at −21℃, a total of 10 coronal sections (50 µm in thickness, between −0.1 and +0.4 mm from bregma) were obtained from each mouse. Four randomly selected sections from each mouse were counterstained with Hoechst 33258 (Santa Cruz Biotechnology, Dallas, TX, USA) for 5 minutes at 24℃ and a coverslip was placed on each slide. The resulting DHE fluorescence was photographed under a laser scanning confocal microscope (LSM700, Zeiss, Munchen, Germany) at 400× and the fluorescence intensity in the high-power field (HPF) was determined using Image analyzer (ImageJ 1.37v, National Institutes of Health, Bethesda, MD, USA). The mean fluorescence intensity was averaged per group.
Neurological deficit scoring
As the first tool for assessing the motor deficits, the neurological deficit scoring (NDS) was employed in accordance with a previous study [26]. In brief, 24-hour post-MCAO/R, motor functions in the mice (n=10 per group) were tested using a scoring scale consisting of 4 points: 0, no neurologic deficit; 1, failure to spread out affected forepaw; 2, unidirectional circling; 3, falling to one side; and 4, no voluntary movement. The NDS was performed by two investigators blinded to the study and the scores were averaged per group.
Grip strength test
The second tool for assessing the motor deficits, the grip strength test, was employed in accordance with a previous study [27]. In brief, after completing the NDS, mice (n=10 per group) were placed on a wire grid attached to custom-made grip test box and allowed to grab the grid with both forepaws before being gently pulled until they released their grip. The maximum force generated was recorded in grams. All tests were conducted in triplicates and the values recorded were averaged per group.
Inverted screen test
The third tool for assessing motor deficits, the inverted screen test, was employed in accordance with a previous study [28]. In brief, after completing the grip strength test, each mouse (n=10 per group) was placed on a custom-made metal grid screen. After placement, the mice were allowed to grip the grid and the screen was inverted to 180° over a cage containing bedding. Latency to fall was recorded up to 180 seconds, and the trials were conducted in triplicates. The values were averaged per group.
Infarct volume measurement
After finishing all the motor function tests, a whole brain of each mouse (n=4 per group) was obtained and cut into eight coronal slices with 1 mm thickness using a brain matrix (Zivic Instruments, Pittsburgh, PA, USA). The sections were stained with 2% of 2,3,5-triphenyltetrazolium chloride (TTC; Sigma, St. Louis, MO, USA) diluted in phosphate-buffered saline (PBS) for 17 minutes at 37℃. The TTC-stained tissues were photographed and ischemic sided brain cortices were immediately isolated and stored at −80℃ until further use in western blotting. The ipsilateral infarct volume was analyzed by an observer blinded to the study with the aid of Image J. To correct the effects of brain edema, the ipsilateral infarcted area in each section was normalized to the contralateral side and expressed as percentage of the contralateral hemisphere. The infarcted areas in each section were summed and averaged per group.
Histologic preparation
After finishing all the motor function tests, the mice (n=6 per group) were anesthetized with an intraperitoneal injection of chloral hydrate (30 mg/kg) and transcardially perfused with 4% paraformaldehyde. Post fixation, each coronal section between −1 and +1 mm from the bregma were trimmed out with the aid of a brain matrix and dehydrated by placing the tissues in an alcohol series. The tissues were then cleared with xylene and embedded in paraffin following which, 5-µm-thick sections were made by a tissue microtome (RM2255, Leica) and mounted on microscope slides. The resulting “sets” for each mouse that contained at least 50 slides, were stored at 24℃ until further use in different experiments, such as, the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay or immunohistochemistry (IHC).
TUNEL assay
The TUNEL assay was conducted by using a commercial kit (DeadEnd, Promega, Madison, WI, USA). According to the manufacturer' protocol, two slides randomly selected from an individual set were assayed and the TUNEL-positive cells of ischemic lesions in at least three randomly chosen HPF were counted under the light microscope (DM4, Leica) at ×400. The counts were averaged per group.
Immunohistochemistry
Each of the two slides randomly selected from an individual set were subjected to an antigen retrieval and endogenous peroxidase quenching step in accordance with a previous study [29]. Next, the slides were incubated overnight with rabbit antibodies against 4-hydroxynonenal (4-HNE), 8-hydroxyl-2′-deoxyguanosine (8-OHdG), and cleaved caspase-3 diluted in PBS at a ratio of 1:200 at 4℃. The slides were then incubated with anti-rabbit IgG diluted in PBS at a ratio of 1:200 for 2 hours at 24℃. All antibodies were purchased from Abcam (Cambridge, UK). Next, the sections were incubated with avidin-biotin complex (Vector, Burlingame, CA, USA) for 1 hour at 24℃. The resulting immunoreactivities were detected using a chromogen, 3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich). After mounting with coverslips, the immunopositive cells in at least three randomly chosen HPF were counted under the light microscope at ×400. The counts were averaged per group.
Western blot
To quantify the level of different antioxidative enzymes, western blot was employed. The lesion-bearing cortices (n=4 per group) that had been stored at −80℃ previously, were homogenized and centrifuged at 120 ×g at 4℃ for 10 minutes. Using a commercially available kit (BCA protein assay kit, Pierce, Rockford, IL, USA), the total protein concentrations were measured in accordance with the manufacturer's protocol. Thereafter, the protein samples were separated by 10% sodium dodecyl sulfate gel and transferred onto a polyvinylidene difluoride membrane (Bio-Rad, Hercules, CA, USA), which was blocked with 5% skim milk diluted in Trisbuffered saline with 0.1% Tween 20 (TBS-T) for 40 minutes at 24℃. The membranes were incubated with rabbit antibodies against superoxide dismutase (SOD), glutathione peroxidase (GPX), heme oxygenase-1 (HO-1), and β-actin diluted in TBS-T at a ratio of 1:1,000 at 4℃ overnight. After washing, the membranes were incubated with anti-rabbit IgG diluted in TBS-T at a ratio of 1:1,000 for 2 hours at 4℃. All antibodies were purchased from Abcam (Abcam). The resulting immunoreactivities were visualized using a chemiluminescence detection system (Immobilon Western, Millipore, Burlington, MA, USA) and photographed using an imaging device (Chemi-doc, Davinch-K, Seoul, Korea). The band intensities were quantified by ImageJ and the results were normalized using β-actin as a house-keeping control. The values were averaged per group.
Statistical analysis
All data were presented as mean±standard error of the mean. Comparisons of the data from the different groups were performed with one-way analysis of variance (ANOVA, PASW Statistics version 18, SPSS Inc., Chicago, IL, USA). Differences with P-values of <0.05 were considered statistically significant. Each “n” value refers to the number of animals.
Results
PSL diminishes fCI-induced infarct volume and behavioral deficits
As shown in TTC-stained tissues (Fig. 2A), there were no visible areas of infarction in the SO group as expected. However, at 24 hours after MCAO/R, the VEH group exhibited marked areas of infarction. The ipsilateral infarct volumes in both the PSL-L and the PSL-H groups were approximately 1/3 of that of the VEH group (13.3±3.6 and 12.9±4.5 vs. 35.5±4.8%, respectively; P<0.001) (Fig. 2B) and the extent of their reduction was not statistically different between these groups. Three different behavioral tests were conducted to assess the role of PSL on fCI-associated motor deficits. First, NDS demonstrated that the motor deficits of the VEH group were prominent (Fig. 3A). However, both the PSL-L and PSL-H groups exhibited significantly improved motor function compared with the VEH group (1.9±0.7 and 1.9±0.7 vs. 2.7±0.9; P<0.01). Second, the grip strength test revealed that grip strength of the VEH group was significantly weakened (P<0.001 vs. SO) (Fig. 3B). Inversely, both the PSL-L and PSL-H groups exhibited significantly higher values in terms of grip strength compared with the VEH group (108.1±18.6 and 139.8±19.6 vs. 76.7±14.6, respectively; P<0.001), and the increase was dose-dependent (P<0.01). Third, the inverted screen test showed that the latencies to fall were significantly reduced in the VEH group compared with the SO group (19.0±14.1 vs. 167.9±15.0; P<0.001) (Fig. 3C). However, the groups treated with PSL remained on the inverted screen longer than VEH group (68.0±14.5 and 140.2±17.4 in PSL-L and -H group, respectively; P<0.001) and this response was dose-dependent (P<0.001). These findings demonstrated that PSL could reduce the infarct volume and improve the motor deficits seen in fCI mice.
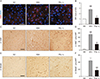
PSL attenuates fCI-induced brain cortical apoptosis
TUNEL staining and IHC detection of cleaved caspase-3, a protein involved in the execution of cellular apoptosis, was employed to assess the extent of cellular apoptosis in brain cortex at 24-hour post-MCAO/R. As shown in Fig. 4A and B, no TUNEL-positive apoptotic cells were detected in brain cortical parenchyma of the SO group as expected. However, presence of TUNEL-positive cells was prominent in the VEH group. In contrast, the number of TUNEL-positive cells was significantly reduced in the groups supplemented with PSL compared with the VEH group (53.5±7.0 and 38.5±13.2 in PSL-L and -H group, respectively, vs. 65.1±6.3; P<0.05 and P<0.01) and this decrease was dose-dependent (P<0.05). Additionally, as shown in Fig. 4C and D, the number of the cleaved caspase-3-immunoreactive cells (indicated by arrows in Fig. 4C) in the ischemic cortex was significantly increased in the VEH group by approximately 6-fold compared with the SO group (59.8±10.6 vs. 9±4.4; P<0.001). However, when compared with the VEH group, the numbers of cleaved caspase-3-immunoreactive cells in the ischemic cortex were markedly decreased in PSL-supplemented groups (39.2±7.8 and 26.4±4.8 in PSL-L and -H group, respectively; P<0.05 and P<0.01) and the inter-group difference was significant (P<0.05). These data suggest that the PSL-induced protection against fCI involves an attenuation of cellular apoptosis.
PSL attenuates fCI-associated oxidative damages
Having established that PSL, even at a low level (20 mg/kg), can reduce the fCI-associated phenotypes, we performed further studies to reveal the underlying mechanism using only the PSL-L group as the treatment group. Given that, a vast majority of phytochemicals are known to exert their beneficial roles on the cellular level via an antioxidant mechanism, we employed in vivo staining with DHE. DHE is a well-known superoxide marker used to identify whether the antioxidant activity is the underlying mechanism of PSL-induced neuroprotection. As shown in Fig. 5A and B, brain ROS accumulation (red fluorescence) was abruptly increased by 2-hour post-fCI insult in the VEH group (2.8±0.5, P<0.001). Conversely, the PSL-L group exhibited a marked reduction of DHE-fluorescence when compared with the VEH group (1.2±0.2, P<0.01). Given that lipid peroxidation and DNA damage are commonly associated with ROS accumulation, we next quantified these mechanisms of damage by employing immunohistochemical detection of 4-HNE and 8-OHdG, which are markers of lipid peroxidation and DNA damage, respectively. As shown in Fig. 5C–F, the results revealed that both 4-HNE- and 8-OHdG-immunopositive stained cells were significantly increased in the brain cortices of the VEH group compared with the SO group (74.0±7.2 for 4-HNE, P<0.001; 44.0±8.5 for 8-OHdG). However, the PSL-L group exhibited a marked reduction in these values when compared with the VEH group (27.8±5.2 for 4-HNE, P<0.001; 17.6%±5.4% for 8-OHdG, P<0.001). Together, these results suggest that PSL can attenuate the fCI-induced ROS accumulation and the subsequent oxidative damage, such as, lipid peroxidation and DNA damage, in brain cells.
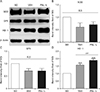
PSL supplement can augment the expression level of an antioxidative enzyme, HO-1
To identify the possible modulation of PSL on brain cellular antioxidative enzyme levels in fCI mice, the protein levels of three enzymes, SOD, GPX, and HO-1, were quantified by western blot technique. As illustrated by the representative band images (Fig. 6A) and the quantification graphs (Fig. 6B–D), the expression levels of SOD and GPX in the ischemic lesion remained unchanged in all the groups. Notably, the levels of HO-1 were significantly increased in the VEH and PSL-L groups compared with the SO group (1.5±0.1 and 1.9±0.1; P<0.001) and the increment was significantly exaggerated in that of PSL-L group (P<0.01). This result suggests that HO-1 can be upregulated by the compensatory response in fCI brain tissue and PSL may further reinforce the upregulation. This finding suggests that upregulation of HO-1, at least in part, may contribute to the neuroprotection of PSL against fCI by reducing oxidative damage.
Discussion
The fCI begins with a permanent or transient insufficiency of cerebral blood flow in the focal brain region. This induces deprivation of oxygen and nutrient supply to brain tissues, leading to an energy failure combined with accumulation of toxic substances [30]. Among these, ROS such as superoxide radicals are known as key molecules that could eventually result in neuronal death [31]. In healthy people, a balance is maintained between ROS production and elimination, which is mediated by the antioxidant enzyme system that is primarily composed of SOD, GPX, and HO-1 [32]. However, the fCI, once initiated, triggers the overproduction of ROS by multiple pathological cascades. The resulting ROS overload in turn threatens neuronal survival owing to their ability to propagate the initial attack on lipid-rich neuronal membranes, causing lipid peroxidation that subsequently results in DNA damage [33]. Accordingly, upregulation of intrinsic neuronal antioxidant enzymes may be an important target for developing fCI therapeutics.
It is well-known that polyphenol compounds, that naturally-occurring plants are enriched with, can combat oxidative stress by upregulation of various endogenous antioxidant enzymes and thus may protect humans against oxidative stress-related pathologies including hypoxia- and inflammation-related diseases [34]. Among the natural plants that are known to trigger this upregulation, PS leaf extract is known to contain several polyphenolic compounds e.g., ellagic acid, gallic acid, and juglone [353637]. As such, it has attracted significant scientific attention [38]. In spite of accumulating evidence that suggest the role of individual compounds of PSL on the activation of antioxidant enzyme-mediated defense system, only limited information is available on the role of PSL itself.
To the best of our knowledge, this study is the first to demonstrate that a PSL supplement can attenuate fCI-induced brain cellular injury via an upregulation of HO-1. This suggests a possible therapeutic mechanism. These findings are consistent with previous studies [3940]. For example, it has been previously reported that neuronal expression of HO-1 is usually detected in rodent brains challenged with anoxic stimuli, including hypoxia or oxidative stress [41]. Furthermore, genetically engineered HO-1-deficient mice exhibit exaggeration in ischemic damage when compared to wild type controls [4243]. Together with the results of this study, these findings suggest that PSL possesses a powerful neuroprotective effect, which may be effective against cerebral ischemia via upregulation of HO-1.
To date, our studies have only focused on identifying the roles of the PSL therapeutic effect against fCI injury in vivo. As such, we have not yet addressed the upstream signal pathway of HO-1 activation that is involved in PSL-mediated neuroprotection. The nuclear factor erythroid-derived 2-related factor 2 (Nrf2) is the master regulator of antioxidative enzymes including HO-1, and Nrf2 is strongly suspected to be one of the most powerful candidates for upstream targeting [44]. Under healthy conditions, a cytosolic Nrf2 is sequestered by binding to Kelch-like ECH-associated protein 1 (Keapl), which inhibits translocation into the nucleus [45]. Several forms of stimuli trigger the conformational change of Keapl that enables the Nrf2/Keap1 complex to release Nrf2, which then translocates to the nucleus and binds to antioxidant-related elements in the promoter regions of antioxidant genes [46]. Additionally, several signaling cascades such as mitogenactivated protein kinase, protein kinase C, and phosphatidylinositol 3-kinase (PI3K/Akt) are involved in the phosphorylation of Nrf2 as well as its nuclear translocation [47]. Hundreds of reports provide evidence for the role of various polyphenols on the activation of Nrf2 [484950]. It is noteworthy that that ellagic acid [51], gallic acid [52], and juglone [53] have been demonstrated to activate the Nrf2/ARE/HO-1 cascade. These reports lead us to assume that the activation of the Nrf2/ARE/HO-1 cascade is mediated by the major components of PSL as described above, that induces a neuroprotective effect during and after fCI. More detailed analysis of the signaling pathways underlying the neuroprotective action of PSL, e.g., the Nrf2/ARE/HO-1 cascade, is important during further investigations.
In conclusion, we revealed that PSL pretreatment can attenuate the infarct volume and improve sensorimotor deficit followed by fCI by exerting antioxidative efficacy. In this regard, we strongly suggest that PSL supplement can be a useful preventive strategy against ischemic stroke.


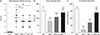

 XML Download
XML Download