

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Global cerebral ischemia is a multifaceted disorder in which brain tissue is subjected to reduced levels of oxygen and glucose due to impairment in blood supply to the entire brain. However, numerous studies have focused on the hippocampus as this important region is one of areas injured [12]. One of the main features of transient global cerebral ischemia is delayed death of pyramidal neurons of the hippocampal CA1 region that occurs hours to days after the insult [2]. After this time-window, neurons can undergo apoptosis/necrosis or survive by maintaining cellular homeostasis [34].
Neuregulin-1 (NRG1) is a family of proteins containing an epidermal growth factor (EGF)-like motif that mediates important functions in the nervous system [567]. NRG1 signaling regulates nerve cell differentiation, neuron migration, neuronal survival, neurite outgrowth, and synaptic activity [68]. NRG1 and its receptor ErbB tyrosine kinases are expressed not only in the developing nervous system, but also in adult brains [910]. Its receptors are tyrosine kinases ErbB2, ErbB3, and ErbB4. ErbB2 and ErbB4, but not ErbB3, can be activated by NRG1. ErbB4 is expressed at synapses [111213]. It has been implicated in synaptic plasticity [101214]. Previous studies have demonstrated that NRG1 can prevent apoptosis of PC12 and SH-SY5Y cells induced by serum deprivation and H2O2 [151617]. Treatment with NRG1 immediately after oxygen-glucose deprivation (OGD) can significantly increase neuronal survival in rat B-35 neuroblastoma cells and cortical neurons [181920]. A number of recent reports have shown that pre-administration of NRG1 could reduce delayed ischemic cortical damage following transient middle cerebral artery occlusion and permanent focal cerebral ischemia in rats [21222324]. However, the involvement of NRG1 pathway for the selective vulnerability of hippocampal neurons to global ischemia has not been fully understood yet. To identify NRG1 effect elicited by ischemic insults, primary hippocampal cultures were subjected to OGD as an in vitro model for global ischemia. Our study demonstrates an important role of ErbB4 in NRG1-mediated neuroprotection, suggesting that endogenous ErbB4 can serve as a novel target for global ischemic therapy.
Materials and Methods
Reagents and antibodies
The NRG1 used in this study was a recombinant polypeptide containing the entire EGF domain of the β-type NRG1 from PROSPEC (East Brunswick, NJ, USA). Antibodies were obtained from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA; ErbB4, sc-283, sc-8050; β-actin, sc-47778; HRP-conjugated anti-rabbit IgG, sc-2004; and HRP-conjugated anti-mouse IgG, sc-2005). AG1478 (Calbiochem, Darmstadt, Germany) and AG879 (Calbiochem) as inhibitors of ErbB4 and ErbB2, respectively, were dissolved in dimethylsulfoxide (DMSO) (Sigma, Louis, MO, USA). The final concentration of DMSO was 0.001% or less when applied to cells.
Primary hippocampal neuronal culture
Primary hippocampal neurons were cultured as described previously [25] . Briefly, the hippocampus was removed from Sprague-Dawley rat embryos (E18) and dissociated by gentle trituration in PBS (Gibco, Carlsbad, CA, USA). Cells were seeded onto poly-L-lysine-coated 12-well plates and cultured in Neurobasal media (Gibco). Experiments were performed at 14 days after seeding (DIV14).
Lactate dehydrogenase release assay
The extent of cell death was assessed by determining the activity level of lactate dehydrogenase (LDH) released into the culture medium. LDH activity was determined using a Cytotox 96 nonradioactive cytotoxicity assay kit (Promega, Madison, WI, USA) according to the manufacturer's instructions. Results are expressed as a percentage of the maximum LDH release obtained upon complete cell lysis.
Terminal deoxynucleotidyl transferase dUTP nick end labeling staining
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining was performed using an in situ cell death detection kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's protocol. Apoptotic primary hippocampal neurons were labeled with TMR red and nuclei were labeled with 10 µM Hoechst dye. The number of apoptotic cells was counted in five random fields using an LSM 510 Meta system (Zeiss LSM 510 laser scanning microscope, Carl Zeiss, Oberkochen, Germany).
Immunofluorescence
Immunostaining of rat hippocampal neurons (DIV14) was performed as described previously [13]. Briefly, neurons were fixed with 4% paraformaldehyde and 4% sucrose in PBS for 20 min. These cells were permeabilized by incubation in PBS containing 1% bovine serum albumin (BSA) and 0.1% Triton X-100 for 30 minutes at room temperature. After washing, neurons were incubated in buffer containing antibodies against mouse ErbB4 (1:100) overnight at 4℃. These neurons were washed and incubated with an appropriate fluorescein isothiocyanate-conjugated secondary antibody for anti-ErbB4. Nuclei were labeled with 10 µM Hoechst dye. Images were visualized using a LSM 510 Meta system (Zeiss LSM 510 laser scanning microscope, Carl Zeiss).
Western blotting
Western blotting was performed as previously described [25]. Samples were resolved using sodium dodecyl sulfate polyacrylamide gel electrophoresis. Proteins were then transferred to nitrocellulose membranes followed by blocking with TBS that contained 5% BSA and 0.05% Tween 20 at room temperature for 1 hour. Membranes were then incubated with anti-ErbB4 (1:1,000, mouse, Santa Cruz Biotechnology) and anti-β-actin (rabbit, 1:5,000, Santa Cruz Biotechnology) antibodies at 4℃ overnight. After washing, blots were developed with horseradish peroxidase-conjugated secondary antibodies and enhanced using a chemiluminescence system (Amersham Pharmacia, California, CA, USA).
Statistical analysis
Data are presented as mean±SEM of three or more independent experiments. For multiple group comparisons, statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Bonferroni's post-hoc test. Student's paired t test was used for comparisons of means between two groups of cells in a single experiment. Values of P<0.05 were considered statistically significant.
Results
OGD induces neurotoxicity and overexpression of ErbB4 receptor in primary hippocampal neurons
OGD is widely used to study mechanisms associated with global ischemic condition in vitro. First, effects of OGD-induced cytotoxicity in primary hippocampal neurons (DIV 14) were examined. Hippocampal neurons will die when oxygen and glucose are deprived of the medium [26]. This can be monitored by LDH that is produced and released into the medium. LDH assays showed that OGD resulted in a significant increase in hippocampal neuronal death. In particular, compared to the control group, OGD significantly increased cell death following exposure for more than 2 hours, although LDH release continued to increase in each subsequent time interval (0, 1, 2, 6, 14 hours) (Fig. 1A). Therefore, in all forthcoming experiments, cells were subjected to OGD for 2 hours. As shown in Fig. 1B and C, OGD for 2 hours induced ErbB4 receptor overexpression in primary hippocampal neurons.
NRG1 attenuates apoptosis induced by OGD
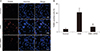
Next, whether NRG1 affected OGD-induced apoptosis in primary hippocampal neurons was examined using TUNEL staining. NRG1 reduced the proportion of apoptotic nuclei in primary hippocampal neurons induced by OGD (Fig. 2A, B). In control and OGD, percentages of apoptotic cells were 3.00%±1.53% and 21.03%±3.22% of total cells, respectively (n=6, P<0.01) (Fig. 2B). Treatment with 5 nM NRG1 for 2 hours significantly reduced the proportion of TUNEL-positive cells (OGD, 21.03%±3.22% vs. OGD+NRG1, 5.33%±1.86%, n=6; P<0.01) (Fig. 2B). The effect of NRG1 on the number of TUNEL-positive cells provided support that NRG1 could attenuate OGD-induced apoptosis in primary hippocampal neurons.
NRG1 pretreatment reduces OGD-induced overexpression of ErbB4 receptor in primary hippocampal neurons
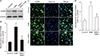
To investigate whether NRG1 could affect OGD-induced increase of ErbB4 receptor expression, cells were pretreated with 5 nM NRG1 followed by 2 hours of OGD. Western blotting was then performed to examine levels of ErbB4 in primary hippocampal neurons. Results confirm that pretreatment with 5 nM NRG1 for 2 hours attenuated the increase of ErbB4 expression induced by OGD (OGD, 2.61±0.34; OGD+NRG1, 1.24±0.26, n=5; P<0.05, P<0.05) (Fig. 3A, B). The immunoreactivity of ErbB4 was then measured via immunofluorescence in primary hippocampal neurons. To examine effects of NRG1 on neurons, cells were pretreated with 5 nM NRG1 and then subjected to OGD 15 minutes later. OGD for 2 hours significantly upregulated ErbB4 expression in comparison with the control (CON, 1.04±0.17; OGD, 2.63±0.31, n=6; P<0.01). Treatment with 5 nM NRG1 for 2 hours attenuated the increase in ErbB4 immunoreactivity induced by OGD (OGD, 3.63±0.31; OGD+NRG1, 1.43±0.21, n=6; P<0.05) (Fig. 3C, D). These results of immunoreactivity were consistent with those of Western blot assay.
ErbB4 is involved in the effect of NRG1 on OGD-induced cell death
NRG1 can activate three ErbB kinases. However, only ErbB2 and ErbB4, but not ErbB3, are catalytically active [27]. To determine which ErbB was involved in the protective effect of NRG1, hippocampal neurons were treated with AG879 (5 µM) and AG1478 (5 µM), known inhibitors of ErbB2 and ErbB4, respectively [24].
However, the protective effect of NRG1was inhibited in neurons after pretreatment with AG1478, suggesting that NRG1 might protect hippocampal neurons by activating ErbB4 kinase. This effect was specific because NRG1 was able to protect neurons after pretreatment with AG879 (Fig. 4). Taken together, these results indicate a role for ErbB4, not ErbB2, in NRG1 neuroprotection.
Discussion
Results of this study demonstrate that ErbB4 is necessary for NRG1-mediated protection of primary hippocampal neurons in in vitro models. When ErbB4 was inhibited, the protective effect of NRG1 on cultured hippocampal neurons after OGD was attenuated. Moreover, expression levels of ErbB4 receptor following OGD were examined. Our results showed a dramatic increase in ErbB4 expression after OGD in cultured neurons. However, pretreatment with NRG1 significantly inhibited OGD-induced abnormal overexpression of ErbB4 receptor in primary hippocampal neurons.
NRG1 and their receptors, ErbB kinases, are continually expressed in mature brain. They are mostly likely to function in neurotransmission and neuroplasticity. NRG1 and ErbB4 are widely expressed in adult brains, including the hippocampus, the cerebellum, and the prefrontal cortex [91328].
Previous studies have indicated that ErbB4 can activate synaptic transmission in the central nervous system. Overexpression of erbB4 enhances AMPA receptor-mediated excitatory postsynaptic currents in rodent CA1 hippocampal pyramidal cells evoked by stimulation of Schaffer collaterals from CA3 [10]. NRG1 enhances γ-aminobutyric acid release from interneurons in the prefrontal cortex [13]. Thus, the neural protective effect of NRG1 might be due to its regulation of synaptic plasticity. In addition, phosphoinositide 3-kinase/Akt signaling pathways might mediate the survival effect of NRG, a critical survival pathway in neurons and most cell types [2930]. Moreover, global cerebral ischemia risk factors promote the production of reactive oxygen species. Oxidative stress can promote inflammation, activate apoptosis signaling pathway and disrupt synaptic activity. NRG1 could attenuate the release of free radicals [17] and protected hippocampal neuronal cells from OGD-induced stress.
On the other hand, high levels of ErbB4 might be pathogenic. Both reduced and increased ErbB4 levels have been associated with schizophrenia [313233]. ErbB4 expression after closed head injury is elevated in neurons. It occurs before its elevation in activated microglia/macrophage [34]. Consistent with this notion is the observation that ErbB4 is upregulated in injured neurons and macrophages/microglia in the penumbra following middle cerebral artery occlusion [35]. Recently, it has been reported that protein levels of ErbB4 are drastically increased in human symptomatic epileptogenic tissues [36].
Several lines of evidence collectively suggest that NRG1 exerts a protective role in neurons against neurotoxic stimuli including ischemic insult and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine [2137]. Previously, our reports have shown that NRG1 signaling plays a neuroprotective role against Swedish mutant and C-terminal fragments of amyloid precursor protein and amyloid beta-peptide Aβ1-42 [383940]. We have also demonstrated that NRG1/ErbB4 can prevent Aβ1-42-induced impairment of long-term potentiation through the phosphoinositide 3-kinase pathway [41]. Taken together, our results suggest that NRG1/EbB4 signaling can protect from oxygen-glucose deprivation insult. Further study is needed to clarify whether pharmacologically targeting ErbB4 in vivo can reduce histological and behavioral abnormalities associated with cerebral ischemia.


 XML Download
XML Download