

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Nerve injury-induced protein (Ninjurin), a cell adhesion molecule, has been reported to be upregulated in the distal and proximal nerves of transected sciatic nerves in rats [1]. Ninjurin family proteins are membrane proteins that have homophilic binding properties [12]. Mammals have two Ninjurin isoforms, Ninjurin-1 and Ninjurin-2, which have similar hydrophobic regions but different adhesion motifs and expression patterns [2].
Ninjurin is expressed widely in embryonic and adult tissues in rats, including the thymus, liver, kidney, and adrenal gland, and is expressed predominantly in epithelial cells where it was thought to be important in normal development and during adulthood [2]. Moreover, apart from its role in nerve regeneration, Ninjurin-1 also considered to involve in thymocyte development [2]. Ninjurin-1 has also been reported to be expressed in endothelial cells and myeloid cells [34]. The function of Ninjurin-1 in early ocular development has been reported to involve mediation of macrophage-induced programmed cell death [3]. In addition, Ninjurin-1 has been reported to exhibit apoptosis-inducing activity in leukocytes, and is cleaved by matrix metalloproteinases, resulting in loss of adhesion during tissue remodeling and pathological conditions [5]. Increased expression of Ninjurin-1 in tissue remodeling mediates cell communication and enhances the entry, migration, and activity of leukocytes, such as monocytes and macrophages in developmental processes [5]. Most of molecules involved in tissue remodeling also play important roles in organogenesis [56]. Therefore, it is important to understand the normal distribution of these molecules, which may be involved in tissue regeneration. In the present study, normal distribution of Ninjurin-1 was investigated in variety of mouse tissues. We report that Ninjurin-1 is predominantly expressed by skin and ileum tissues while least was associated with cerebrum. Because Ninjurin-1 is thought to be involved in tissue remodeling, its normal distribution with current findings further suggests the possible role of Ninjurin-1 in tissue homeostasis.
Materials and Methods
Animals
Male C57BL/6 mice (6–8 weeks old) were obtained from Central Lab (Seocho-gu, Seoul, Korea) and were acclimated for 1 week before use. All experimental procedures were conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals of Jeju National University, Jeju City, Korea (2018-0029). The animal protocols conformed to current international laws and the policies of the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, 1985, revised 1996). Every effort was made to minimize the number of animals used and to minimize their suffering.
Tissue preparation and histological examination
Experimental animals were sacrificed under ether anesthesia, and the cerebrum, sciatic nerve, spleen, lung, liver, pancreas, kidney, testis, skin, stomach, ileum, and colon were dissected and homogenized in lysis buffer for protein analysis. Pieces of tissues were fixed in 4% (v/v) paraformaldehyde in phosphate-buffered saline (PBS; pH 7.4) for 48 hours. Subsequently, the sections (5 µm thick) were used for hematoxylin and eosin staining and for immunohistochemistry.
Western blot analysis
Western blotting was performed as described previously [7]. Briefly, protein lysates (2 µg/µl) were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, and the resolved proteins were immunoblotted onto nitrocellulose membranes (Whatman, Dassel, Germany). The residual binding sites on the membranes were incubated with mouse monoclonal anti-Ninjurin-1 antibody (1:1,000 dilution, BD Transduction Laboratories, CA, USA). The membranes were washed three times in Tris-buffered saline containing 0.1% Tween-20 and then incubated with horseradish peroxidase-conjugated anti-mouse IgG (Vector Laboratories, Burlingame, CA, USA). Immunoreactive bands were visualized using a BS ECL Plus kit (Biosesang, Seongnam, Korea) according to the manufacturer's protocol. After imaging, the membrane was stripped and reprobed using mouse anti-β-actin antibody (Sigma-Aldrich, St. Louis, MO, USA). Immunoblot signals were detected by Fusion Solo 6X (Vilber Lourmat, Collegien, France) and analyzed using FUSION® software. The expression values were normalized to that of β-actin. The cerebrum band in each panel was used as a standard for normalization, and the relative density of each band was calculated. The results are presented as the mean±standard error.
Immunohistochemistry
Immunohistochemistry was performed using a commercial avidin-biotin complex kit (Vector Laboratories), as described in our previous study [7]. Briefly, deparaffinized sections were treated with 0.01 M citrate buffer (pH 6.0) in a microwave oven for 2 minutes, and then treated with 0.3% hydrogen peroxide in methyl alcohol for 20 minutes to block endogenous peroxidase activity, followed by incubation with 10% (v/v) normal horse serum in PBS. The samples were incubated with primary antibody, mouse anti-Ninjurin-1, for 1 hour at room temperature. After three washes in PBS, the peroxidase reaction was developed using a diaminobenzidine substrate kit (Vector Laboratories). The sections were counterstained with hematoxylin, dehydrated, and cleared with xylene before finally being mounted.
Results
Expression of Ninjurin-1 in various tissues
Western blot analyses were performed to evaluate the expression levels of Ninjurin-1 in various mouse tissues. The results showed that Ninjurin-1 was least expressed in the cerebrum, while it was abundant in skin and ileum tissues (Fig. 1A). Other mouse tissues, including sciatic nerve, lung, liver, pancreas, kidney, testis, stomach, and colon, showed moderate Ninjurin-1 expression compared to that of the cerebrum. Based on semi-quantitative analysis (Fig. 1B), the level of Ninjurin-1 was particularly high (about 10-folds) in skin and ileum compared with that in the cerebrum, which was arbitrarily set to 1, and moderate (about 5-folds) in sciatic nerve, spleen, lung, liver, pancreas, kidney, testis, stomach, and colon. These findings indicated that the level of Ninjurin-1 varied among the investigated organs.
Dynamic cellular expression of Ninjurin-1 in different organs
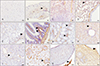
Immunohistochemical analysis showed differential Ninjurin-1 immunoreactivity in the cerebrum, sciatic nerve, spleen, lung, liver, pancreas, kidney, testis, skin, stomach, ileum, and colon (Table 1, Fig. 2). In the cerebrum (Fig. 2A), Ninjurin-1 immunoreactivity was not detected in neurons and was only detected in some glial cells (Fig. 2A). In the sciatic nerve (Fig. 2B), positive immunoreactivity of Ninjurin-1 was associated with some axons and with Schwann cells (Fig. 2B, arrowhead).
In the spleen (Fig. 2C), cells in the red pulp and some white pulp cells were immunopositive for Ninjurin-1. In the lung (Fig. 2D), immunoreactivity for Ninjurin-1 was detected in alveoli, alveolar macrophages (Fig. 2D, arrowhead), and adventitia in bronchioles (Fig. 2D).
In the stomach (Fig. 2E), parietal cells were negative for anti-Ninjurin-1 reactivity. Instead, immunoreaction was detected in some chief cells (Fig. 2E, arrowhead), areas of the submucosa and tunica muscularis.
In the ileum (Fig. 2F), some cells in lamina propria (Fig. 2F, arrowheads) were immunopositive for Ninjurin-1. Moreover, positive immunoreactivity was detected in the tunica muscularis and tunica serosa. In the colon (Fig. 2G), the lamina propria (Fig. 2G, arrowheads) was immunopositive, whereas epithelial cells were immunonegative for Ninjurin-1. Both the tunica serosa and tunica muscularis were immunopositive for Ninjurin-1.
In the liver (Fig. 2H), some hepatocytes and sinusoids showed a positive immunoreaction along with immunopositive Kupffer cells (Fig. 2H, arrowhead) and vascular endothelial cells. In the pancreas (Fig. 2I), both acinar cells were not immunopositive for Ninjurin-1, but the interlobular ducts and pancreatic islets (Fig. 2I, arrowhead) were immunopositive.
In the kidney (Fig. 2J), some cells in the glomeruli (Fig. 2J, arrowhead), including glomerular capillary cells, mesangial cells, and the basal surface of proximal tubules, were immunopositive for Ninjurin-1. In the testis (Fig. 2K), spermatogenic cells and interstitial cells (Fig. 2K, arrowhead) were immunopositive for Ninjurin-1. In the skin (Fig. 2L), sebaceous cells and keratinocytes (Fig. 2L, arrowhead) were immunopositive for Ninjurin-1. Moreover, collagen fibers and some hair follicles were immunopositive for Ninjurin-1.
Discussion
It is becoming apparent that Ninjurin-1 plays multiple roles in tissue regeneration and pathogenesis [18]. Most of our findings are consistent with the previous studies, which showed differential expression of ninjurin mRNAs in embryonic and adult rat tissues [12]. To our knowledge, this is the first study to show the expression of Ninjurin-1 in most of the tissues we investigated including lung, pancreas, stomach and intestines.
Low level of Ninjurin-1 in the cerebrum, and its' cellular localization were matched with previous studies [1349]. However, under inflammatory conditions, Ninjurin-1 is upregulated in the brain with dynamic cellular expression levels [410]. Ninjurin-1 was first discovered in peripheral nerve injury in rats, where low level was associated with uninjured sciatic nerve [1]. Ninjurin-1, also reported to contribute mainly to peripheral nervous system development and repair [2]. Similarly, in the present study, taken together, Ninjurin-1 expression in the peripheral nervous system was higher than that of the central nervous system.
A previous study showed Ninjurin expression in adult rat spleen [1]. Ninjurin-1 was upregulated in the spleen after Yersinia pestis infection in mice [11] and in septic mice [12]. Depending on the present study we propose that Ninjurin-1 plays a role in T and B cell homeostasis in the normal spleen. It has been reported that the lung is a major site of T cell proliferation and assume migratory properties before entering the inflamed central nervous system [13]. As Ninjurin-1 is also a well-known adhesion molecule for immune cell migration [4] its' expression in normal lung tissue may act as a signaling molecule for T cell trafficking under inflammatory conditions.
Expression of Ninjurin-1 was detected in some hepatocytes and liver sinusoids in consistent with previous studies [12]. Compared to wild-type mice, Ninjurin-1-deficient mice are prone to systemic inflammation in the liver [14]. Up-regulation of Ninjurin was observed in hepatocellular carcinomas than in normal or fetal liver tissues [15]. Ninjurin-1 may play a role in hepatocyte regeneration in normal and pathological liver.
Even though Ninjurin-1 was moderately expressed in the pancreas based on Western blotting, immunohistochemical staining revealed that acinar cells were negative for Ninjurin-1. Instead, the interlobular duct was immunopositive for Ninjurin-1. Moreover, Ninjurin-1-deficient mice are prone to pancreatic insulitis with mixed T and B cell infiltration surrounding the pancreatic islets [14]. Therefore, expression of Ninjurin-1 in normal pancreas may be indicative of a self-protective mechanism in pancreas.
Expression of Ninjurin-1 in urogenital organs is less reported. In a previous study, Ninjurin was detected in podocytes and mesangial cells of the glomeruli, but not in other renal cell types [2]. Similarly, in the present study, Ninjurin-1 was expressed in some cells of the glomeruli, particularly mesangial cells, mesangium in kidney and spermatogenic cells and interstitial cells of the testis. But the mechanism involving Ninjurin-1 in normal urogenital organs has not been investigated.
Ninjurin-1 expression in keratinocytes and fibroblasts was reported both in normal and some pathological conditions [10]. In the current findings, Ninjurin-1 immunoreaction was mostly associated with connective tissues of the dermis, some keratinocytes, and almost all sebaceous cells. Current finding is supported by the previous reports of Ninjurin-1 immunoreactivity in epithelium of adult rat skin tissue [1]. Expression of Ninjurin-1 mRNA was increased in keratinocytes after exposure to X-irradiation together with genes related to apoptosis [10]. But, a contrast finding reported that Ninjurin-1-heterozygous mice were more prone to skin lesions than wild-type mice [14]. However, Ninjurin-1 in normal skin tissue may be regulated together with other genes to maintain skin stability and quick cellular regeneration following damages.
Ninjurin-1 expression in the ileum was approximately 10-folds higher than cerebrum. It has been suggested that Ninjurin-1 overexpression helps in suppression of colon cancer development [16]. Therefore, Ninjurin-1 expression in normal gastrointestinal tissues may help to fence against oncogenic factors.
Collectively, we postulate that Ninjurin-1 is expressed in all organs and may play important roles in organs with high or moderate expression levels, including the spleen, lung, liver, kidney, testes, skin, ileum, and colon. However, further experiments are remained in order to unravel the specific molecular mechanisms involving Ninjurin-1 in each organ system. Because, we cannot exclude the possibility that the role of Ninjurin-1 is concealed in organs with low expression, where the cell phenotype of Ninjurin-1 may be less distinctive.


 XML Download
XML Download