

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
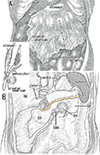
The transverse colon in adults is located below the stomach and behind the greater omentum (Fig. 1A), with a root of the transverse mesocolon running longitudinally across the anterior aspect of the pancreatic body or along the inferior margin (Fig. 1B). During embryonic gastric rotation along the longitudinal axis [12], the greater omentum originates from the dorsal mesogastrium, the site of development of major parts of the pancreas. Because the omentum and transverse mesocolon are composed of double layered peritoneum, a laminar arrangement in the upper abdomen was hypothesized (Fig. 1 inset). This type of logical arrangement may be difficult to show in dissection of adult cadavers, but separation of the greater omentum from the mesocolon, or omentectomy, is regarded as a classical procedure in surgery [3]. In Japan, resection of regional lymphatics may include separation of the transverse mesocolon not only from the greater omentum but also from the omental bursa or lesser sac [4]. The ability to separate the peritoneal fusion in adults may enable demonstration of the logical peritoneal lamination in fetuses. Examination of sections from human fetuses showed histologic evidence of the fusion fascia behind the duodenopancreas, a fusion used in a routine surgical maneuver for pancreaticoduodenectomy (Kocher's maneuver or retropancreatic pancreaticoduodenal mobilization) [5]. That study, however, did not show macroscopic views before the preparation of histological sections.
Determining the topographical anatomy of the stomach and intestines in fetuses before, during and after physiological umbilical herniation is of major interest in embryology [67]. Excellent 3D studies have demonstrated alterations in the topographical anatomy of the transverse colon [89]. However, since all these studies used specimens earlier than Carnegie stage 22 or 23 (gestational age [GA] 7–8 weeks), little is known about when and how the transverse mesocolon fuses with the greater omentum. A study using serial sagittal sections without 3D reconstruction showed that the descending colon entered into the herniation sac in 16 of 27 embryos and fetuses of GA 6–9 weeks [10]. Therefore, despite the anatomy of the greater omentum and transverse mesocolon being well known, inter-individual differences likely occur. Previous macroscopic studies, however, did not include descriptions of peritoneal structures (e.g., Rigoard et al. [11]), likely because no or few studies combined gross anatomy with histological observations. This study, using sections from midterm and late-stage human fetuses (crown-rump length [CRL], 32–325 mm), was designed to demonstrate (1) the development of fusion between the greater omentum and transverse mesocolon and (2) the fetal topographical relationship between the stomach and transverse colon. During dissections of late-stage specimens for histological preparation, histologic characteristics were compared with macroscopic views.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in 2013). The study included 45 midterm fetuses of CRL 32–110 mm (GA 8–15 weeks) and 17 late-stage fetuses of CRL 250–328 mm (GA 30–38 weeks). All specimens were part of the large collection kept at the Embryology Institute of the Universidad Complutense, Madrid, and originated from miscarriages and ectopic pregnancies at the Department of Obstetrics of the University. The midterm specimens had been sectioned serially, 10 sagittally and 35 horizontally; most sections were stained with hematoxylin and eosin (H&E), with others stained with Masson trichrome, azan, orange G or silver stain. In contrast, after macroscopic observations during dissection, the abdomens of the late-stage fetuses were sectioned horizontally at 100–200-µm intervals and stained with H&E. During dissection of the late-stage fetuses, photographs were taken with a Pentax K-1 camera equipped with a 50–100 mm zoom lens. Histological samples were observed and most photographs taken with a Nikon Eclipse 80, whereas photographs at ultra-low magnification (objective lens less than ×1) were taken with a high-grade flat scanner and translucent illumination (Epson scanner GTX970, Java, Indonesia). The study protocol was approved by the Ethics Committee of the Universidad Complutense (B08/374).
Results
Observations of midterm fetuses
Sagittal sections of 10 specimens (CRL, 32–78 mm; GA, 8–12 weeks) and horizontal sections of 35 specimens (CRL, 36–110 mm; GA, 8–15 weeks) were evaluated. Physiological umbilical herniation was seen in four of these specimens, of CRLs 32, 35, 38, and 45 mm, one sectioned sagittally and three sectioned horizontally.
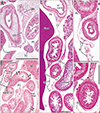
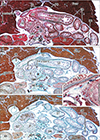
The transverse colon in all 10 sagittally sectioned specimens (Fig. 2) was located below the stomach and behind the greater omentum. One of these specimens with physiological herniation (CRL, 32 mm) showed a separation between the transverse mesocolon and the greater omentum (Fig. 2A), whereas the transverse mesocolon in the other nine sagittally sectioned specimens was fused to the greater omentum around and above the colon (Fig. 2B, C). However, the fusion plane was not identified at higher magnification (Fig. 2D, E). In two of the 10 sagittally sectioned specimens, the continuous mesentery of the ascending, transverse and descending colons was free of retro-abdominal structures, except for the pancreas, with this mesentery showing a reverse-U shaped arrangement surrounding the jejunum and ileum (Fig. 2B). The double layers of the peritoneum were often difficult to identify at sites without vessels or fat in the greater omentum. Similarly, the peritoneum was found to be a thin layer of mesothelium on the pancreas, stomach and colon.
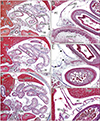
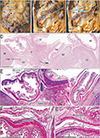
The transverse colon in most of the horizontally sectioned specimens was present in sections below the pylorus or antrum of the stomach. In these sections, the greater omentum was usually cut between the liver and colon, but it was sometimes folded and pushed by the jejunum to the liver left lobe. Notably, in six specimens, of CRLs 48, 55, 85, 90, 97, and 110 mm (GA, 10–15 weeks), part of the transverse colon was attached to the anterior aspect of the gastric antrum and pylorus (Figs. 3, 4, 5). Although most of these colons were simply attached, the colon and stomach in one specimen, of CRL 97 mm, were connected by fibrous tissue (Fig. 5 inset). The transverse colon in these six specimens with prepyloric colon ran medially along the liver right lobe from the hepatic flexure, extended along the gastric greater curvature, and passed leftward and posteriorly to reach the splenic flexure. The final half of the course was situated between parts of the jejunum (Fig. 3A, B) or along the liver left lobe (Fig. 4C). The greater omentum extended along the liver left lobe, but was fused, in whole or in part, with the transverse mesocolon (Fig. 3E). The transverse mesocolon was also fused with the duodenopancreas (Fig. 4E–G) as well as with the renal fascia (Fig. 4H). The bag-like double layers of peritoneum were observed along the omentum (Fig. 3D, E). In addition, the duodenal loop always provided a C-loop, with its descending portion directed inferiorly. Thus, both the second and fourth portions were contained in a single horizontal section (Figs. 4D, 5C).
Observations of late-stage fetuses
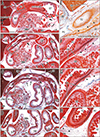
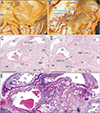
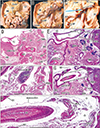
All 17 late-stage fetuses (GA 30–38 weeks) were macroscopically observed in a step-by-step manner during dissection for preparation of histological specimens (Figs. 6, 7, 8, 9). In 15 specimens, the greater omentum extended inferiorly while shifting leftward (Fig. 6A, B), likely because the dilated ileum was located in the right half of the abdominal cavity. In one specimen (Fig. 6C), the greater omentum shifted rightward, possibly due to the dilated jejunum; and in one specimen (Fig. 7A), the greater omentum extended evenly and widely, covering almost the entire ileum and jejunum. Beneath the transparent omentum, the transverse colon extended along the left-right axis below the stomach. However, the right sided part of the transverse colon appeared to push the pylorus and/or the superior portion of the duodenum upward (Figs. 7A, 8A). When the transverse colon was reflected upward, a fan-like arrangement of branches of the middle colic artery and vein was observed in the transverse mesocolon (Fig. 7B). A cut line of the transverse mesocolon ran longitudinally across the pancreatic body (Figs. 7C, 8B). The mesocolon in 14 of the 17 specimens was attached to the right aspect of the superior or descending portion of the duodenum. However, the presence of bulky fat associated with the omentum and/or mesocolon made it difficult to determine macroscopically whether the colon was actually attached to the stomach (see below).
Histologic examination of five late-stage specimens showed that the transverse colon was attached to the right lower aspect of the gastric antrum in three specimens (Figs. 7D, 9D) and to the anterior aspect of the pylorus in two (Fig. 8C). The greater omentum or both the omentum and mesocolon were sandwiched by or wedged between the stomach and colon were observed at the attachment sites (Figs. 7F, 9F, H), depending on the arteries contained in the tissue samples. However, the omentum and mesocolon were not fused but separated by a narrow space. Likewise, a narrow space was seen at the attachment site of the transverse colon to the duodenum (Figs. 7G, 8E, 9H), but arteries of the colon appeared not to be involved at the interface. Histologic sections of a single specimen strongly suggested that the transverse colon pushed the superior portion of the duodenum in an upward direction (Fig. 9E, G). Finally, all duodenal loops were C-shaped, but the descending portion was often directed posteriorly. Thus, in these specimens, almost the entire duodenal loop was contained in a single horizontal section (Figs. 8D, 9D). In addition, when compared with our previous data [12], a lymphatic tissue development was not evident along the greater omentum and mesocolon as well as a fusion plane of the peritoneum along the duodenum and pancreas.
Discussion
The present study demonstrated that the transverse mesocolon fused with the greater omentum at GA 8–9 weeks and that subsequently these peritoneal structures could not be distinguished in the common mesentery immediately after repackaging from physiological herniation. However, at mid-term, we observed variants in the topographical anatomy of the transverse colon and stomach, including (1) a prepyloric transverse colon (GA 10–15 weeks), and (2) a transverse colon pushing the superior portion of the duodenum and pylorus upward (GA 10–15 weeks and 30–38 weeks). These variations seemed to modify the initial fusion between the mesocolon and greater omentum because the stomach is encapsulated by peritoneal laminae other than the transverse colon (Fig. 1). Actually, the colon was likely to tightly fuse with the stomach although the former maintained, in whole or in part, fusion with the greater omentum. We also found variations in the peritoneal configuration in late-stage fetuses: the stomach and transverse colon were likely to sandwich the greater omentum and/or the mesocolon (GA 30–38 weeks). Moreover, therein, the omentum and mesocolon were not fused but separated by a narrow space. Consequently, the initial fusion between the greater omentum and mesocolon was likely to be modified, changed or even broken in the later development and growth.
Those variants in the topographical anatomy of the transverse colon and stomach seemed to be caused by the more rapid growth of the colon than the stomach within the limited volume of the abdominal cavity. With the growth of this cavity, the prepyloric colon returned to its normal position, with the incidence of prepyloric colons being much lower in late-stage than in midterm fetuses. Similarly, a colon located behind and below the stomach was likely to push the stomach upward. Detailed morphometry using magnetic resonance images of 377 embryos showed deflection of the stomach during the embryonic period [13], a deflection that may be caused by upward growth of the transverse colon. Jeong et al. [14] described that, in late-stage human fetuses, a fusion plane of the transverse mesocolon with the other peritoneal structures is not simple but provides irregular and rough surfaces. Before the present study, we had considered this irregularity results from the secondarily degeneration of lymphatics after a great mass of lymphatic tissues appears along the peritoneal fusion in the upper abdomen at 18–20 weeks [12]. However, according to their figures, the temporally increased lymphatics is not evident along a fusion plane between the greater omentum and transverse mesocolon.
Surgical maneuvers to separate the fused omentum and mesocolon (see the Introduction) may be performed simply along an avascular space, depending on the supplying arteries, i.e., the middle colic artery to the transverse mesocolon and the epiploic arteries to the greater omentum. Our findings did not exclude the possibility that surgeons find a secondary loosening or separation of the peritoneal fusion due to fetal topographical changes among the stomach and transverse colon. Another possibility also likely that, in adults with some pathological conditions involving increased lymphatic flow, the mesocolon and omentum may separate along their actual fusion plane in development. During the same period as the transverse mesocolon is fused with the greater omentum, a reversed-U shaped arrangement of the continuous mesentery of the entire colon [10] may also fuse with the renal fascia. Similar to a separation of the greater omentum from the mesocolon, a surgical separation of the mesocolon from the renal fascia is also a basic surgical maneuver [1516], but there may be a “trick” for the easy separation in fetal development as seen in the omentum-mesocolon fusion in the present study. Consequently, a logical lamination of the peritoneum seemed not to simply connect with the surgical application.
The greater omentum also exhibited variations in the direction of extension, with the most striking feature being the rarity of usual wide omenta in fetuses. We had regarded a left or right shift of the omentum as resulting from inflammation in mesenteric lymph nodes and omental milky spots after birth. Indeed, folding, fusion and adhesion of the greater omentum are often seen in adult cadavers [17]. However, some of these variations were likely established in fetuses even without abnormal rotation of their intestines. The location of dilated parts of the intestine as well as individual variations in upper abdominal anatomy, such as prepyloric colon, seemed to determine the morphology of the greater omentum. The leftward growth of the great curvature of the stomach has also accelerated the left shift of the omentum. Finally, we stated about an almost horizontal duodenal loop at late stage of the duodenum in contrast to the usual C-loop in the frontal plane: this temporal horizontal loop might result from differences in growth rates between portions of the duodenum. Overall, after repackaging of physiological herniation, topographical anatomy in the upper abdomen seemed not to be stable but continue to change.

 XML Download
XML Download